Факторы активации клеток врожденного иммунитета

Распознавание PAMP подготавливает клетки врожденного иммунитета к выполнению их основной функции — удалению чужеродных агентов из внутренней среды организма. При этом происходит экспрессия ряда генов, появляются отсутствовавшие на покоящихся клетках молекулы, участвующие в выполнение клетками своих эффекторных функций. Переход клетки в состояние, обеспечивающее выполнение ею своих функций, обозначают термином «активация».
При активации происходит экспрессия определенных наборов инду- цибельных генов. Различия эффекторных функций разных типов клеток обусловлены экспрессией различных наборов генов. При активации для большинства клеток врожденного иммунитета характерны: появление новых молекул на поверхности клеток (в частности молекул адгезии и разнообразных рецепторов), секреция цитокинов и других гуморальных продуктов, усиление метаболизма.
Источник активации клеток врожденного иммунитета — связывание рецепторами своих лигандов с последующей передачей в клетку активационного сигнала, траснформируемого в сигнал, индуцирующий экспрессию генов. Для индукции генов необходимо образование в клетке ядерных (транскрипционных) факторов, обладающих сродством к определенным последовательностям ДНК и связывающихся с регуляторным (промоторным) участком соответствующих генов. В покоящихся клетках наборы транскрипционных факторов, необходимых для индукции этих генов, отсутствуют. Появление факторов транскрипции достигается разными путями: активацией предсуществующих неактивных факторов с их перемещением в ядро, синтезом этих факторов de novo или разрушением их инактиваторов. Факторы, необходимые для реализации этих процессов, должны быть в свою очередь индуцированы при активации клеток. Именно поэтому между мембранным рецептором, поставляющим активационный сигнал, и генами с их регуляторными участками расположена цепь передаточных (сигнальных) молекул. В процесс активации вовлечено несколько сигнальных путей, приводящих к образованию разных транскрипционных факторов. В состав внутриклеточных сигнальных путей входят ферменты — киназы (фосфорилируют белки или липиды, переводя их в активное состояние) и адапторные белки (передают промежуточные продукты активации между звеньями сигнальной цепи.
Основные факторы, вызвающие активацию клеток врожденного иммунитета — PAMP, узнаваемые патогенраспознающими рецепторами (в первую очередь — TLR). Передача сигнала по сигнальным путям проиллюстрирована на рис. 2.12 и 2.13. При связывании PAMP с мембранным TLR возникают конформационные изменения внеклеточной части рецептора, передающиеся на внутриклеточный домен TIR. В передаче активационного сигнала от TIR-домена ^R участвует несколько адапторных белков (MyD88, TIRAP, TRIF, TRAM), активирующих 2 основных сигнальных пути — MyD88-зависимый и TRIF-зависимый. MyD88-зависимый путь участвует в передаче сигнала от всех ^R, кроме TLR-3, использущего TRIF-зависимый путь. Передача сигнала от TLR-4 происходит с участием обоих сигнальных путей. MyD88 (при участии TIRAP) играет роль «мостика» между активным димером TLR и первой сигнальной киназой — серин- треониновой киназой IRAK-4. Активированная IRAK-4 запускает каскад реакций активации сигнальных ферментов: киназы IRAK-1, убиквитин- лигазы TRAF6 и киназы ТАК1.
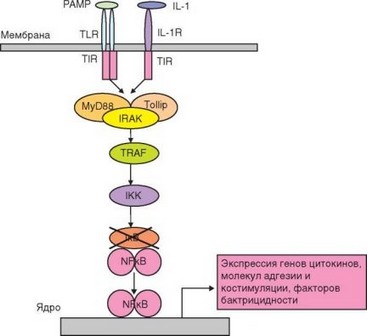
Рис. 2.12. Основной сигнальный путь, активируемый при связывании лигандов Toll-подобными рецепторами и приводящий к экспрессии транскрипционного фактора NF-kB и активации провоспалительных генов
На этом этапе передача сигнала может идти по двум путям. Один из них — активация тирозинфосфатазы IKK. Активация IKK происходит также при поступлении сигналов от эндолизосомального TLR-3 при посредстве адапторного белка TRIF и киназы RIP1. Основная мишень IKK — IkB (ингибирующая цепь неактивного комплекса, содержащего транскрипционный фактор NF-kB). Фосфорилирование IkB вызывает ее связывание с убиквитином, после чего она подвергается расщеплению в протеасоме. Освобожденный от IkB комплекс содержит активный димер NF-kB, миг- рирущий в ядро и связывающийся с промоторными участками многих провоспалительных генов (цитокинов, молекул адгезии, бактерицидных пептидов, ферментов и т.д.). Таким образом достигается главная цель активации — превращение клеток в эффекторы, обеспечивающие развитие воспалительной реакции и реализацию защитных функций врожденного иммунитета.
Второй путь передачи сигнала, раздваивающийся на уровне киназы ТАК1, состоит в активации MAP-каскада (MAP — от mitogen-activated proteinkinase) — серии последовательных активаций серинтреониновых
протеинкиназ от MAP-киназ 3-го (ТАК1) до 1-го уровня (JNK и p38). MAP-киназы 1-го уровня обеспечивают образование транскрипционного фактора АР-1 (Activation protein 1). АР-1 участвует в активации многочисленных генов, имеющих отношение к развитию не только воспаления, но и адаптивного иммунного ответа (см. раздел 3.5.2.1).
Передача сигнала от TLR, локализованных в эндолизосомах, происходит другим способом. От TLR-7, TLR-8, TLR-9 сигнал передается с участием адапторного белка MyD88 путем последовательной активации IRAK4, IRAK1, TRAF6 и ТАК1. Следующие за этим пути передачи сигнала также расходятся (рассмотрены выше). Они приводят к образованию транскрипционных факторов NF-kB и AP-1. Однако, в отличие от мембранных TLR, при передаче сигнала от эндолизосомальных TLR формируется дополнительная сигнальная ветвь. При формировании околорецепторного мультимолекулярного комплекса, включающего MyD88, IRAK4, IRAK1, TRAF3,
TRAF6, неактивный IRF7 (IRF — Interferone-responding factor) и некоторые другие факторы, происходит активация IRF7. Активированный IRF7 мигрирует в ядро и, соединяясь с последовательностью ISRE (Interferon-stimulated response element), играет роль транскрипционного фактора, ответственного за «включение» гена интерферона a (IFNa) (см. рис. 2.13).
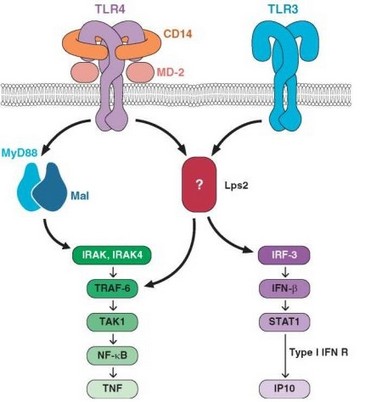
Передача сигнала от рецепторов TLR-3 и TLR-4 (при его экспрессии в эндолизосомах) происходит иным путем, но приводит к тем же результатам. Прежде всего в сигнальной цепи отсутствует MyD88. Роль первого адаптор- ного белка при этом играет TRIF (для TLR-4 — также TRAM). TRIF имеет участки связывания с белками RIP1 и TRAF3, инициирующими 2 пути передачи сигнала. Один из них состоит в активации киназы RIP1, активации IKK и формированию транскрипционного фактора NF-kB. Активация убиквитин-лигазы TRAF3 приводит (через промежуточную стадию с участием факторов TBK1 и IKKe) к активации фактора IRF3. Этот транскрипционный фактор индуцирует экспрессию генов интерферонов, причем в большей степени IFNp, чем IFNa.
Суммируя рассмотренные выше данные о сигнальных путях (см. рис. 2.13), можно констатировать образование 4 транскрипционных факторов, участвующих в развитии воспаления и проявлений врожденного иммунитета: NFkB (ключевой транскрипционный фактор провоспалительных генов), АР-1 (транскрипционный фактор для включения различных иммунологически значимых генов), IRF7 и IRF3 (транскрипционные факторы, ответственные за включение генов интерферонов — соответственно IFNa и IFNp). При этом мембранные TLR (TLR-5 и функциональный комплекс TLR-1/TLR-2/TLR-6) участвуют в активации NF-kB и АР-1, эндолизосомальные TLR (TLR-7, TLR-8 и TLR-9) ответственны за включение этих факторов и дополнительно IRF7, а мембранные рецепторы (TLR-3 и TLR-4) — за включение NF-kB и АР-1 и дополнительно IRF3. Таким образом, TLR, распознающие внеклеточные патогены, передают сигналы, приводящие к экспрессии провоспалительных генов, а TLR, распознающие внутриклеточные патогены (в частности вирусы), помимо провоспалительных, индуцируют гены интерферонов, обеспечивающих противовирусную защиту.
После взаимодействия мембранных TLR с лигандом происходит их интернализация и отделение от фактора MyD88. Это служит одним из факторов, обусловливающих временную «неотвечаемость» на повторное действие того же агента — толерантность, проходящую только через 2—3 сут. Реакция, развивающаяся при связывании TLR-4 с лигандом, отличается от описанной выше: TLR-4 интернализуется и теряет связь с MуD88, но сохраняет связь с фактором TRIF, что обусловливает его функционирование в составе эндолизосомы, о чем говорилось выше.
Изучение передачи сигнала от цитозольных паттернраспознающих рецепторов семейства NLR—NOD1/2 показало, что по результатам она сходна с передачей сигнала от мембранных TLR. Связывание с NLR их лигандов (мурамилпептидов) приводит к активации (при участии фактора RICK) комплекса IKK с последующим формированием фактора NF-kB и активацией каскада MAP-киназ c образованием транскрипционного фактора АР-1. При этом активации генов интерферонов не происходит.
Как уже сказано, к основным генам, активируемым под влиянием NF-kB, относят гены провоспалительных цитокинов. При экспрессии генов семейства IL-1 для синтеза функционально активного продукта (прежде всего IL-1P) необходим процессинг синтезированной молекулы- предшественницы, состоящий в ее расщеплении каспазой 1. В процессинге задействованы рецепторы NALP (цитозольные рецепторы семейства NLR), формирующие вместе с другими факторами и прокаспазой (все они содержат домен CARD) надмолекулярный комплекс инфламмосому, в которой и происходит активация каспазы 1.
Из приведенных выше данных следует, что PAMP-распознающие рецепторы, относящиеся к TLR и NLR, — главные факторы активации миелоид- ных клеток, задействованных в реакциях врожденного иммунитета. Другие паттернраспознающие рецепторы ответственны за выполнение функций, не требующих активации клеток, однако они могут участвовать в этом процессе в качестве корецепторов. Пример таких рецепторов — молекулы адгезии интегрины. Они связаны с тирозинкиназами и молекулами, имеющими активационные мотивы ITAM. Таким образом, интегрины способны активировать факторы, общие для нескольких путей активации ^м. 2.3.1.2), что способствует образованию транскрипционных факторов NF-kB и AP-1. Один из С-лектиновых рецепторов — дектин-1 имеет в своей цитоплазматической части последовательность ITAM, участвующую в передаче активационных сигналов. Связывание дектина-1 с р-глюканами дрожжевых форм грибов приводит к индукции провоспалительных генов, в том числе циклооксигеназы-2 и цитокинов, в частности TNFa, IL-6, IL-23, что определяет роль дектина-1 в защите от грибковой инфекции. Другие лектиновые рецепторы самостоятельно не участвуют в активации клеток, хотя и могут способствовать TLR-зависимой активации клеток.
Анализ сигнальных путей, активируемых при связывании провоспалительных цитокинов, выявляет очень высокую степень их сходства с MyD88-зависимой передачей сигнала (см. раздел 2.5.5.3). Для IL-1 эти пути идентичны сигнальным путям, запускаемым при связывании TLR, поскольку внутриклеточная часть рецептора для IL-1 представляет TIR-домен (что отражено в названии этого домена — Toll/IL-1 receptor and resistance domain). При образовании TNFa в передачу сигнала вовлечены факторы TRAF2 и TRAF3, что обусловливает наличие перекрестов с сигнальными путями TLR и TNFR. Результат сходства этих путей передачи сигнала — явление, называемое амплификацией ответа на PAMP. Оно заключается в том, что эффект, достигаемый при прямом действии патогенов и их продуктов в очаге инфицирования, дистантно воспроизводится полностью и даже в значительно большем масштабе за счет действия провоспалительных цитокинов на клетки врожденного иммунитета. Таким образом, амплифи- цирующие факторы —продукты PAMP-индуцированной активации.
Выше детально рассмотрен классический путь активации клеток врожденного иммунитета (прежде всего макрофагов), на котором основано вовлечение этих клеток в иммунную защиту и воспалительные реакции. Однако резидентным макрофагам и дендритным клеткам свойственны и другие формы ответа, направленные на осуществление гомеостатических функций, регенерацию тканей, а также ограничение воспалительных процессов. В их основе лежат иные пусковые факторы и сигнальные механизмы. Альтернативные пути активации миелоидных клеток на примере макрофагов представлены в табл. 2.12.
Таблица 2.12. Варианты активации макрофагов под влиянием различных сигналов
| Показатель | Варианты активации | ||
| Классический | Альтернативный | Активация II типа | |
| Активирующий сигнал | INFy + TNF (TLR) | IL-4, IL-13 | IgG-комплексы + TLR, CD40 или CD44 |
| Секреторные продукты | t TNF, t IL-12, IL-1, IL-6 | t IL-1RA IL-10 | t IL-10, i IL-12, TNF, IL-1, IL-6 |
| Поверхностные маркеры | t MHC-II, t CD86, i Маннозо-связы- вающий рецептор, i Fcy-RII | t Маннозосвязывающий рецептор, t Рецепторы-мусорщики, t CD23, i CD14 | t MHC II класса, t CD86 |
| Ферменты | t NO-синтаза, i Аргиназа | t Аргиназа, i NO-синтаза | i NO-синтаза |
| Секретируемые хемокины | IP-10 (CXCL10), MIP-1a (CCL3), MCP-1 (CCL2), | PARC-1 (CCL18), MDC (CCL22), TARC (CCL17) | Неизвестно |
| Биологические эффекты | Повышение бактерицидной и тумороцидной активности, презентация антигена | Ускорение регенерации, подавление пролиферации Т-клеток, уменьшение бактери- цидности | Противовоспалительная активность, усиление антителопродукции |
Таким образом, при действии разнообразных чужеродных (патогены) и эндогенных (цитокины) лигандов на рецепторы клеток врожденного иммунитета запускается весь комплекс процессов, необходимых для осуществления защиты — эндоцитоз (поглощение) чужеродных агентов и активация, приводящая к мобилизации защитных механизмов и секреции активных факторов защиты.
Источник
Активация врожденного иммунитета. Стадия активации врожденного иммунитетаВрожденная иммунная защита первично нацелена на распознавание и элиминацию чужеродных структур. Инфекционные агенты располагают многими факторами вирулентности, воздействие которых может быть разрушительным для защитных механизмов врожденного иммунитета. Организм хозяина выработал уникальные стратегии распознавания патогенных структур микроорганизмов, процессинга их и презентации антигенов Т-лимфоцитам. Активация врожденного иммунитета, обеспечивающая оптимальное развитие специализированных эффекторных механизмов, может быть концептуально разделена на две стадии: 1 — фаза детекции, в течение которой комбинация поверхностных рецепторов распознает присутствие чужеродных структур при вторжении микрорганизмов, и 2 — фаза трансмиссии этой сенсорной информации на язык, понятный для клеток адаптивной иммунной системы, например, через продукцию хемокинов и цитокинов. Универсальная стратегия распознавания микроорганизмов эффекторами врожденного иммунитета основана на детекции доменных структур, которые называются патоген-ассоциированными молекулярными образами — PAMPs (pathogen-associated mollecular patterns), а распознающие их рецепторы врожденной иммунной системы — образраспознающими рецепторами — PRRs (pattern-recognition receptors). PAMPs — консервативные (неспецифические, невариабельные) структуры микроорганизмов общие для разных патогенов, их нет у млекопитающих, поэтому они распознаются иммунной системой как «чужое» с помощью паттерн-распознающих рецепторов (PRRs), включая семейство сигнальных Toll-подобных рецепторов (TRL). Существуют еще и альтернативные рецепторы врожденного иммунитета, такие как нуклеотид-связывающий олигомеризующийся домен (nucleotide-binding oligomerization domain — NOD), локализующийся внутриклеточно. NOD распознают РАМР независимо от участия TLR, активируют ядерный фактор KB (NF-kB) и усиливают продукцию IFN-y.
Эти эволюционно законсервированные рецепторы распознают PAMPs, включающие различные компоненты бактериальной стенки, среди которых наиболее известные — это липополисахариды, пептидогликан, липотейхоевые кислоты, маннаны, флагеллин, бактериальная ДНК, вирусные двуспиральные РНК, глюканы. До недавнего времени рассматривалось, что PAMPs обычно экспрессируются на поверхностях микробов и отсутствуют на клетках организма хозяина, однако в последнее время появляются работы, демонстрирующие наличие паттернов хозяина, распознаваемых рецепторами этого класса. В настоящее время известно два типа рецепторов врожденного иммунитета, обеспечивающих информирование организма о проникновении патогена. К первому типу относятся растворимые рецепторы для патогенов: C1q комплемента, ЛПС-связывающий белок, маннозосвязывающий лектин, С-рективный белок. Второй тип рецепторов для патогенов — рецепторы, экспрессирующиеся клетками врожденного иммунитета. Закодированные в геноме, рецепторы врожденной иммунной системы имеют ряд отличий от рецепторов лимфоцитов. Они экспресси-руются на эффекторных клетках врожденной иммунной системы, осуществляющих первую линию защиты, а также на профессиональных антигенпрезентирующих клетках (макрофагах, дендритных клетках и В-лимфоцитах). — Также рекомендуем «Варианты образраспознающих рецепторов PRRs. Toll-подобные рецепторы» Оглавление темы «Врожденный иммунитет»:
|
Источник
Врожденный иммунитет — это первая линия обороны иммунной системы, которая включает предсуществующие механизмы защиты, всегда готовые к быстрой, стереотипной обороне. Рецепторы клеток врождённого иммунитета генетически закодированы и неизменны в течение жизни; на поверхности микроорганизмов они распознают структуры жизненно важных для микробов молекул, которые не могут быть изменены в результате одной мутации. Клетки врожденного иммунитета одного и того же типа имеют одинаковый набор рецепторов. Они находят и убивают болезнетворные микроорганизмы и одновременно активируют адаптивный иммунный ответ. Врожденный (неспецифический) иммунитет обеспечивает защиту с умеренной эффективностью в течение нескольких дней, пока не активируется адаптивный иммунитет. Адаптивный иммунитет для активации требует от нескольких дней до недели. Адаптивный иммунитет специфический, его составляющие научены ответу на точные молекулярные структуры. Для создания своих рецепторов адаптивный иммунитет использует перегруппировку генов (реаранжировку), и пролиферацию клеток со специфическими рецепторами путем формирования клонов. Адаптивный иммунитет имеет иммунологическую память. Адаптивная иммунная система — дополнение к эволюционно более древней врождённой иммунной системе, обеспечивающее специфичность распознавания и память.
Частью врожденного иммунитета являются барьеры организма. Они включают в себя эпителиальные слои, слизь для предотвращения прилипания микробов и антимикробные пептиды. Эти пептиды включают α — дефензины нейтрофилов, β — дефензины эпителиальных клеток и гистатины слюны.
В эффекторных механизмах иммунной системы (воспаление и реакция острой фазы) задействованы опсонизация, фагоцитоз, внутриклеточное уничтожение и секреция цитокинов. Бактерии имеют механизмами уклонения от эффекторных механизмов защиты, эффекторные механизмы иммунитета имеют ограниченное действие против вирусов.
Клеточное звено врожденного иммунитета представлено разными видами лейкоцитов: нейтрофилы составляют 50-70% лейкоцитов, лимфоциты 20-35%, моноциты 3-7%, эозинофилы 1-3% и базофилы 0-1%.
")
- Нейтрофилы и макрофаги являются наиболее важными фагоцитирующими клетками. Они образуются из полипотентных гемопоэтических стволовых клеток и дальнейшего дифференцирования миелоидного — предшественника (гранулоцито-моноцито колониеобразующей клетки).
 Нейтрофилы и макрофаги обычно используют одни и те же механизмы уничтожения чужого, но нейтрофилы живут около суток, активируются при остром воспалении, и уничтожают только бактерии с использованием активных форм кислорода. Макрофаги, в отличие от нейтрофилов, живут недели, действуют при хроническом воспалении, атакуют многие микроорганизмы, презентируют антиген, секретируют множество цитокинов и используют оксид азота как реактивную форму кислорода.
Нейтрофилы и макрофаги обычно используют одни и те же механизмы уничтожения чужого, но нейтрофилы живут около суток, активируются при остром воспалении, и уничтожают только бактерии с использованием активных форм кислорода. Макрофаги, в отличие от нейтрофилов, живут недели, действуют при хроническом воспалении, атакуют многие микроорганизмы, презентируют антиген, секретируют множество цитокинов и используют оксид азота как реактивную форму кислорода.
Работа нейтрофилов – фагоцитировать чужое. Нейтрофилы широко представлены циркуляции и тканях, и они очень мобильны и поэтому обычно первыми реагируют на возбудителя. В азурофильных гранулах нейтрофилы содержат гидролитические ферменты, дефензины и миелопероксидазу. Другие гранулы переносят рецепторы для комплемента, адгезии и цитокинов и готовы к экзоцитозу получив сигнал. Незрелые нейтрофилов еще не имеют характерного ядра полиморфноядерных клеток (зрелых нейтрофилов), ядро в виде палочки. Для распознавания и связывания мишеней нейтрофилы используют в основном Fc рецепторы и рецепторы комплемента, рецепторы распознавания образов.
")
Макрофаги фагоцитируют патогены и презентируют антиген. Они могут что-то фагоцитировать, и освобождать антииммунные (толерогенные) сигналы, не секретировать сигналы, или секретировать проиммунные (иммуногенные) сигналы. В крови они
циркулируют как моноциты в течение суток, затем мигрируют в ткани, где дифференцируются в макрофаги (больше цитоплазмы, гранул и складчатая мембрана). Макрофаги имеют рецепторы к комплементу, Fc и рецепторы распознавания образов.- Макрофаги являются важнейшими регуляторами как адаптивного, так и врожденного иммунного ответ.
- Естественны киллеры (ЕK) являются важными эффекторными лимфоцитами врожденного иммунитета, которые проявляют цитолитическую активность против различных аллогенных внутриклеточных мишеней в неспецифических, контакт-зависимых, не фагоцитарных процессах, которые не требует предварительной сенсибилизации антигеном. ЕК клетки имеют несколько свойств обычных цитотоксических Т-клеток (ЦЛТ), в том числе аналогичные механизмы цитолиза. Цитолитическая активность ЕК опосредована формированием пор в клетке-мишени с последующей секрецией в мишень белков, таких как гранзимы и перфорин, сериновые протеазы и другие. Их цитотоксическая активность позитивно регулируется с помощью IL-2, IL-15 и интерферонов, и негативно простагландинами и TGF-β.
- Естественные киллеры Т клетки (ЕКТ) – представляют собой различные линии Т-клеток, которые экспрессируют инвариантный Т-клеточный рецептор αβ (TCR αβ) и имеют на поверхности ряд маркеров, общих с ЕК. ЕКТ клетки рестриктированы по неполиморфной CD1d молекуле и активируются гликолипидами антигенов, представленными CD1d. Для идентификации мышиных и человеческих ЕKT может быть использованы CD160 и Vα24Jα18 соответственно
- Интраэпителиальные γδ Т-клетки (освобождают провоспалительные цитокины),
- B-1 клетки (синтез неспецифических «естественных» антитела).
Врожденный иммунитет активируется паттерн-распознающими рецепторами (PRRs), которые узнают паттерны патоген ассоциированных молекул (РАМРs, подобно ЛПС, CpG DNA, fMet, dsRNA и др.).
Паттерны патоген ассоциированных молекул должны быть экспрессированы на патогене, но отсутствовать у хозяина. Они имеют тенденцию к структурной инвариантности для группы возбудителей, и жизненно необходимы патогенам. Это полисахариды / нуклеотиды, но не белки.
Паттерн распознающие рецепторы (PRRs) являются генетически закодированной линией и не подвержены реаранжировке.
Взаимодействие паттерн распознающих рецепторов с патером патоген ассоциированных молекул инициирует внутриклеточные сигнальные каскады, которые обычно заканчиваются передачей сигнала в ядро и синтезом провоспалительных цитокинов.
Источник