Iga и клеточный иммунитет

Циркулирующие иммунные комплексы (ЦИК) представляют собой комплексы, состоящие из антител, антигена и компонентов комплемента. Образование ЦИК – нормальный механизм защиты организма, но иногда, вследствие избыточного накопления, ЦИК сохраняются в течение длительного времени и откладываются в различных тканях и органах, приводя к их поражению (при системной красной волчанке (СКВ)).
Уровень ЦИК в крови – показатель развития различных воспалительных процессов в организме и показатель активности течения аутоиммунных заболеваний.
Показания
· диагностика и наблюдение за течением аутоиммунных и ревматоидных заболеваний (диффузных болезней соединительной ткани, системных васкулитов. СКВ, болезни Крона, аутоиммунного гепатита, коллагенозов и др.);
· диагностика и наблюдение за течением хронических инфекционных заболеваний (ВИЧ-инфекции, подострого инфекционного эндокардита и др. вирусных, бактериальных и грибковых заболеваний);
· наблюдение за течением хронических воспалительных заболеваний почек (гломерулонефрита);
· в комплексе с другими исследованиями для диагностики иммунодефицитных состояний (входит в исследование иммунного статуса)
Методика
Определение ЦИК в сыворотке крови осуществляется методом иммунного турбидиметрического анализа.
С3 и С4-компоненты комлемента
Белки компонента комплемена синтезируются в печени и в норме присутствуют в плазме в неактивной форме. Пусковыми механизмами активации системы комплемента является проникновение в организм бактерий и вирусов, грибов, простейших.
Показания:
диагностика болезней иммунных комплексов — системная красная волчанка (СКВ), генерализованные васкулиты, гломерулонефрит, энтеропатии (болезнь Крона, неспецифический язвенный колит), аутоиммунная гемолитическая анемия.
При рецидивирующих инфекциях: стрептококковом и стафилококковом сепсисе, бактериальных и вирусных менингитах, паразитарных инвазиях.
Мониторинг лечения болезней иммунных комплексов— СКВ, аутоиммунный тиреоидит, ювенильный дерматомиозит.
Диагностика наследственного дефицита компонентов комплемента -ангионевротический отек или болезнь Квинке, дефицит С3 при рекуррентных инфекциях, дефицит С4 при СКВ.
Методика
Определение С3 и С4 – компонентов комплемента осуществляется турбодиметрическим методом на биохимическом анализаторе «АRCHITECT с8000»
Иммунный статус (субпопуляционное определение абсолютного и относительного количества лимфоцитов CD3+, CD4+, CD8+, CD19+, естественных киллеров CD16+, общих IgA, IgM, IgG, ЦИК, подсчет ИРИ)
Для оценки клеточного иммунитета определяются его основные параметры (относительное содержание клеток в крови): зрелые Т-клетки, Т-хелперы, Т-супрессоры /цитотоксические, В-клетки, NK-клетки (естественные киллеры) и иммуно-регуляторный индекс (ИРИ) – соотношение Т-хелперов и Т-супрессоров/цитотоксических.
Показания
· выявление первичного и вторичного иммунодефицита,
· контроль за проведением иммуностимулирующей терапии.
Методика
Определение основных параметров клеточного иммунитета проводится методом проточной цитометрии с использованием реагентов «Лаборатория КОНСТАНТА» на проточном цитометре «Partec CyFlow».
Кроме показателей клеточного иммунитета, в иммунный статус входят показатели гуморального иммунитета: общие IgA, IgM, IgG, и ЦИК.
Подготовка
Взятие крови предпочтительно проводить утром натощак, после 8 – 14 часов ночного периода голодания (воду пить можно), допустимо днем через 4 часа после легкого приема пищи. Накануне исследования необходимо исключить психоэмоциональные и физические нагрузки (спортивные тренировки), прием алкоголя, за час до исследования – курение. Отказаться от приема лекарственных препаратов: даназол, гидралазин, фенитоин, антибактериальных препаратов пенициллинового ряда, метилдопа, цимстидин, пероральные контрацептивы.
Источник
Иммуноглобулин А. Секреторный IgA в иммунитете слизистых оболочек.
Известно, что степень защиты от локальных вирусных инфекций респираторного и желудочно-кишечного трактов прежде всего зависит от содержания в организме специфического секреторного IgA, а не от наличия сывороточного IgG к энтеропатогенным или пневмотропным вирусам.
Стабильная структура, выраженный аффинитет к поверхности слизистых оболочек, преобладающее содержание в секрете молочной железы обусловливают биологическую роль секреторного IgA в защите организма от неблагоприятного воздействия различных патогенных агентов, в том числе вирусов.
IgA синтезируется в димерной форме в клетках lamina propria и после связывания с иммуноглобулиновым рецептором, синтезированным в эпителиальных клетках, транспортируется на поверхность слизистой оболочки. В момент выхода IgA в просвет кишечника рецептор частично расщепляется, в результате чего в составе IgA остается фрагмент рецептора, который называют секреторным компонентом.
Таким образом, секреторный IgA является продуктом кооперации двух типов клеток — плазматических и эпителиальных.
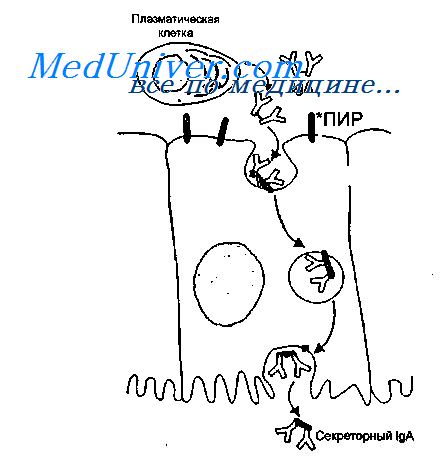
Секреторный IgA образуется не только в димерной, но и в тетрамерной форме, что усиливает его вируснейтрализующую способность. Секреторный компонент предохраняет IgA от расщепления протеолитическими ферментами, что обусловливает его значительные преимущества перед антителами других классов. Секреторный IgA нейтрализует вирус не только в просвете кишечника, но и при транспортировке его внутрь клетки. Димер IgA может нейтрализовать вирус в подслизистой оболочке кишечника, а затем, связавшись с рецептором, транспортировать его в просвет кишечника.
Димерные иммуноглобулины класса A (US IgA), соединенные посредством j-цепи в единую структуру с секреторным компонентом, представляют собой уникальный пример эволюционной адаптации иммуноглобулинов на слизистых покровах для эффективного функционирования в условиях постоянного воздействия антигенов различной природы.
Исходя из способности к локальному синтезу молекул секреторного компонента, а также димерной формы IgA-местно расположенными плазматическими клетками, ткани слизистых покровов можно разделить на три основных класса.
В слизистых оболочках индуктивная и продуктивная фазы гуморального иммунного ответа разобщены пространственно в большей степени, чем в других отделах иммунной системы.
Клеточные и гуморальные превращения, связанные с появлением IgA во внешних секретах, представлены на рисунке.
В молоко и, по-видимому, в другие внешние секреты молекулы IgA поступают из двух главных источников. Большая часть IgA, выделяемого со слюной, секретами слезной и молочной желез, а также пищеварительного и респираторного трактов, образуется плазматическими клетками. Однако IgA, обнаруживаемые в различных внешних секретах, могут иметь и системное происхождение. Продуцируясь клетками слизистых оболочек одних органов, они поступают в кровь и переносятся в слизистые покровы других органов.
Антиген после проникновения внутрь пейеровой бляшки активирует Т- и В-лимфоциты, которые по лимфатическому протоку поступают в мезентериальный лимфатический узел, а затем в кровь, селезенку, снова в кровь и селективно локализуются в лимфатических образованиях всех слизистых оболочек и экзокринных секреторных железах — молочных, слюнных и слезных. При этом Т-лимфоциты локализуются преимущественно между эпителиальными клетками слизистой оболочки, обеспечивая клеточный иммунный ответ, а В-лимфоциты — в lamina propria, где они дифференцируются в плазматические клетки и синтезируют IgA.
Около 90% плазматических клеток в lamina propria продуцируют IgA, в то время как в лимфатических узлах доля таких клеток составляет всего 2—5%.
Уникальна в этом отношении роль печени. Имеются убедительные данные, свидетельствующие о том, что гепатоциты избирательно связывают и в последующем транспортируют IgA в желчь, тем самым усиливая систему секреторного IgA кишечника.
Возможной функцией печени также является удаление комплексов антиген — IgA из циркулирующей крови с желчью в кишечник. Изучение противовирусного иммунитета убедительно продемонстрировало, что степень защиты респираторного и желудочно-кишечного трактов от локальных вирусных инфекций прямо коррелирует с уровнем специфических секреторных IgA, а не с уровнем антител сыворотки крови. В основе противовирусного действия секреторного IgA лежит инактивация вируса.
— Также рекомендуем «Кишечник и иммунитет. Роль кишечника в работе иммунной системы человека.»
Оглавление темы «Патогенез и механизмы противовирусной защиты организма.»:
1. Иммунная система слизистых оболочек. Слизистая оболочка и вирусная инфекция.
2. Строение иммунной системы слизистых. Концепция общей иммунной системы слизистых оболочек.
3. Иммуноглобулин А. Секреторный IgA в иммунитете слизистых оболочек.
4. Кишечник и иммунитет. Роль кишечника в работе иммунной системы человека.
5. Респираторные органы и вирусная инфекция. Роль дыхательной системы в противовирусном иммунитете.
6. Молочная железа и вирусная инфекция. Роль молочной железы в противовирусном иммунитете.
7. Особенности иммунитета при вирусных инфекциях. Патогенез противовирусного иммунного ответа.
8. Разрушение эффекторных клеток иммунитета и макрофагов вирусами. Устранение действия цитокинов вирусами.
9. Индукция иммунологической толерантности вирусами. Иммуносупрессия при вирусной инфекции.
10. Гуморальный противовирусный иммуннитет. Клеточный противовирусный иммунный ответ.
Источник
Существуют две ветви приобретенного иммунитета с разным составом участников и различным предназначением, но имеющие одну общую цель — устранение антигена. Как мы увидим в дальнейшем, эти две ветви взаимодействуют друг с другом, чтобы достичь конечной цели — устранения антигена.
Из этих двух направлений приобретенного иммунного ответа одно определяется участием в основном В-клеток и циркулирующих антител, в форме так называемого гуморального иммунитета (термин «гуморальный» ранее использовали для определения жидких сред организма). Другое направление определяется участием Т-клеток, которые, как мы указывали ранее, не синтезируют антител, но синтезируют и высвобождают различные цитокины, действующие на другие клетки. В связи с этим данный вид приобретенного иммунного ответа называется клеточным или клеточно-опосредованным иммунитетом.
Гуморальный иммунитет
Гуморальный иммунитет определяется участием сывороточных антител, которые являются белками, секретируемыми В-клеточным звеном иммунной системы. Первоначально после связывания антигенов со специфическими молекулами мембранного иммуноглобулина (Ig) (В-клеточные рецепторы; В cell receptors — BCR) В-клетки активируются для секреции антител, которые экспрессируются этими клетками. По имеющимся оценкам, каждая В-клетка экспрессирует примерно 105 BCR совершенно одинаковой специфичности.
После связывания антигена В-клетка получает сигналы на производство секретируемой формы того иммуноглобулина, который ранее был представлен в мембранной форме. Процесс инициации полномасштабной реакции с участием антител направлен на удаление антигена из организма. Антитела представляют собой гетерогенную смесь сывороточных глобулинов, которые обладают способностью самостоятельно связываться со специфичными антигенами. Все сывороточные глобулины со свойствами антител относят к иммуноглобулинам.
Все молекулы иммуноглобулинов имеют общие структурные свойства, которые позволяют им: 1) распознавать и специфически связываться с уникальными элементами структуры антигена (т.е. эпитопами); 2) выполнять общую биологическую функцию после соединения с антигеном. В основном, каждая молекула иммуноглобулина состоит из двух идентичных легких (L) и двух тяжелых (Н) цепей, связанных дисульфидными мостиками. Получающаяся в результате структура показана на рис. 1.2.
Рис. 1.2. Типичная молекула антитела, состоящая из двух тяжелых (Н) и двух легких (L) цепей. Выделены антигенсвязывающие участки
Часть молекулы, которая связывается с антигеном, является зоной, состоящей из терминальных участков аминокислотных последовательностей как на L-, так и на Н-цепях. Таким образом, каждая молекула иммуноглобулина является симметричной и способна связываться с двумя идентичными эпитопами, имеющимися на одной молекуле антигена или на разных молекулах.
Кроме различий между участками, связывающими антиген, у разных молекул иммуноглобулина имеются и другие различия, наиболее важные из которых касаются Н-цепей. Существует пять основных классов Н-цепей (называемых у, μ, α, ε и δ).
На основании различий в Н-цепях молекулы иммуноглобулина были разделены на пять основных классов: IgG, IgM, IgA, IgE и IgD, каждый из которых характеризуется уникальными биологическими свойствами. Например, IgG является единственным классом иммуноглобулинов, пересекающим плацентарный барьер и передающим материнский иммунитет плоду, в то время как IgA — основной иммуноглобулин, обнаруживаемый в таких секретах желез, как слеза или слюна.
Важно отметить, что антитела всех пяти классов могут обладать совершенно одинаковой специфичностью по отношению к антигену (антигенсвязывающие участки), сохраняя в то же время различные функциональные (биологические эффекторные) свойства.
Связь между антигеном и антителом нековалентная, она зависит от множества относительно слабых сил, таких как водородные связи, вандерваальсовы силы и гидрофобные взаимодействия. Поскольку эти силы слабы, для успешного связывания антигена с антителом требуется очень близкий контакт на ограниченном участке, наподобие контакта ключа и замка.
Другим важным элементом гуморального иммунитета является система комплемента. Реакция между антигеном и антителом активирует комплемент, который составляют ряд сывороточных ферментов, что приводит или к лизису мишени, или усиливает фагоцитоз (поглощение антигена) клетками-фагоцитами. Активация комплемента также приводит к привлечению полиморфно-ядерных (ПМЯ) клеток, обладающих высокой способностью к фагоцитозу и являющихся частью врожденной иммунной системы. Эти события обеспечивают максимально эффективный ответ гуморальной ветви иммунитета на вторжение чужеродных агентов.
Клеточно-опосредованный иммунитет
Антигенспецифичная ветвь клеточно-опосредованного иммунитета задействует Т-лимфоциты (рис. 1.3). В отличие от В-клеток, вырабатывающих растворимые антитела, которые циркулируют для связывания соответствующих специфичных антигенов, каждая Т-клетка, несущая множество идентичных антигенных рецепторов, называемых TCR (около 105 на клетку), сама направляется непосредственно к месту, где на АПК экспрессируется антиген, и взаимодействует с ней в близком (непосредственно межклеточном) контакте.

Рис. 1.3. Рецепторы для антигена, экспрессируемые как трасмембранные молекулы на В- и Т-лимфоцитах
Существует несколько различающихся по фенотипу субпопуляций Т-клеток, каждая из которых может обладать одинаковой специфичностью по отношению к антигенной детерминанте (эпитопу), но при этом выполнять различные функции. В данном случае можно провести аналогию с разными классами молекул иммуноглобулинов, которые обладают одинаковой специфичностью, но различными биологическими функциями. Имеются две субпопуляции Т-клеток: Т-клетки-хелперы (Тн-клетки), которые экспрессируют молекулы CD4, и цитотоксические Т-клетки (Тс-клетки), которые экспрессируют молекулы CD8 на своей поверхности.
Разным субпопуляциям Тн-клеток приписывают различные функции.
- Взаимодействие с В-клетками для увеличения продукции антител. Такие Т-клетки действуют путем высвобождения цитокинов, которые обеспечивают подачу различных активирующих сигналов В-клеткам. Как указывалось ранее, цитокины являются растворимыми веществами или медиаторами, высвобождаемыми клетками; такие медиаторы, высвобождаемые лимфоцитами, называются лимфокинами. Группе цитокинов с низкой молекулярной массой дали название хемокины. Они, как указывается далее, участвуют в воспалительной реакции.
- Участие в реакциях воспаления. После активации определенная субпопуляция Т-клеток высвобождает цитокины, индуцируя миграцию и активацию моноцитов и макрофагов, что приводит к возникновению так называемых воспалительных реакций гиперчувствительности замедленного типа. Эту субпопуляцию Т-клеток, участвующих в реакции гиперчувствительности замедленного типа (ГЗТ), иногда называют Тгзт или просто Тн.
- Цитотоксические эффекты. Т-клетки особой субпопуляции становятся цитотоксическими клетками-киллерами, которые при контакте со своей мишенью способны нанести удар, ведущий к гибели клетки-мишени. Эти Т-клетки называют цитотоксическими Т-клетками (Тс). В отличие от Тн-клеток они экспрессируют молекулы CD8 на своих мембранах и поэтому называются СD8+-клетками.
- Регуляторные эффекты. Хелперные Т-клетки могут быть разделены на две различные функциональные подгруппы в соответствии с цитокинами, которые они высвобождают. Как вы узнаете из следующих глав, эти субпопуляции (Тн1 и Тн2) обладают различными регуляторными свойствами, которые передаются посредством высвобождаемых ими цитокинов. Более того, Тн1 -клетки могут негативно перекрестно влиять на Тн2-клетки, и наоборот. У другой популяции регуляторных или Т-клеток-супрессоров отмечается коэкспрессия CD4 и CD25 (CD25 является α-цепью рецептора интелейкина-2. Регуляторная активность этих СD4+/СD25+-клеток и их роль в активном подавлении аутоиммунитета обсуждается в гл. 12.
- Эффекты цитокинов. Т-клетки и другие клетки иммунной системы (например, макрофаги) оказывают различное воздействие на многие клетки, лимфоидные и нелимфоидные, посредством разных цитокинов, которые они высвобождают. Таким образом, прямо или косвенно Т-клетки связываются и взаимодействуют с множеством типов клеток.
В результате многолетних иммунологических исследований было установлено, что клетки, активированные антигеном, проявляют целый ряд эффекторных способностей. Однако только за последние несколько десятилетий иммунологи стали осознавать всю сложность событий, которые происходят при активации клеток антигеном и при их взаимодействии с другими клетками. Мы теперь знаем, что простой контакт Т-клеточного рецептора с антигеном недостаточен для активации клетки.
В действительности для активации антигенспецифичной Т-клетки должны быть даны по крайней мере два сигнала. Первый сигнал обеспечивается связыванием Т-клеточного рецептора с антигеном, который должен быть соответствующим образом презентирован АПК. Второй сигнал определяется участием костимуляторов, среди которых имеются определенные цитокины, такие как IL-1, IL-4, IL-6, и поверхностные молекулы, экспрессированные на АПК, такие как CD40 и CD86.
В последнее время под термином «костимулятор» стали подразумевать и другие стимулы, например продукты жизнедеятельности микроорганизмов (инфекционные, чужеродные) и поврежденная ткань («гипотеза опасности» П. Матзингера (P. Matzinger)), которые будут усиливать первый сигнал, если он относительно слаб. Как только Т-клетки получают достаточно четкий сигнал для активации, происходит ряд событий, и активированная клетка синтезирует и высвобождает цитокины. В свою очередь эти цитокины контактируют с определенными рецепторами на различных клетках и воздействуют на эти клетки.
Хотя обе, гуморальная и клеточная, ветви иммунного ответа рассматриваются как самостоятельные и отличные друг от друга компоненты, важно понимать, что реакция на любой специфический патоген может предусматривать сложное взаимодействие между ними, а также участие элементов врожденного иммунитета. Все это нацелено на обеспечение достижения максимально возможного выживания организма за счет удаления антигена и, как мы увидим далее, защиты организма от аутоиммунного ответа на собственные структуры.
Проявление разнообразия в иммунном ответе
Последние достижения в иммунологических исследованиях обусловлены союзом молекулярной биологии и иммунологии. Благодаря тому что клеточная иммунология смогла выявить на клеточном уровне суть многочисленных и различных по спектру реакций, а также природу процессов, позволяющих достичь уникальной специфичности, появилось множество соображений относительно реальных генетических механизмов, которые позволяют всем этим специфичностям стать частью репертуара у каждого представителя данного вида.
Вкратце эти соображения таковы:
- По различным подсчетам число специфичных антигенов, к которым может возникать иммунный ответ, способно достигать 106—107.
- Если каждый специфичный ответ, как антительный, так и Т-клеточный, определяется одним геном, означает ли это, что каждому индивидууму потребуется более 107 генов (один на каждое специфичное антитело)? Каким образом этот массив ДНК передается неповрежденным от индивида к индивиду?
На этот вопрос позволили ответить новаторские изыскания, проведенные С.Тонегавой (S.Tonegawa) (лауреат Нобелевской премии) и Ф.Ледером (Ph.Leder), в которых были использованы методы молекулярной биологии. Эти исследователи описали уникальный генетический механизм, с помощью которого иммунологические рецепторы, экспрессированные на В-клетках и отличающиеся огромным разнообразием, могут создаваться на базе относительно небольшого количества ДНК, предназначенного для этой цели.
Природа создала технологию генных рекомбинаций, при которой белок может кодироваться молекулой ДНК, составленной из набора рекомбинируемых (переставляемых) мини-генов, которые и составляют полный ген. На основе небольшого набора таких мини-генов, способных свободно комбинироваться для создания целого гена, можно получить огромный репертуар специфичностей, используя ограниченное число генных фрагментов.
Первоначально этот механизм был призван объяснить существование огромного разнообразия антител, которые не только секретируются В-клетками, но также фактически составляют антиген-или эпитопспецифичные рецепторы В-клеток. Впоследствии было установлено, что подобные механизмы отвечают и за разнообразие антигенспецифичных Т-клеточных рецепторов (TCR).
Достаточно сказать, что существование различных методов молекулярной биологии, позволяющих не только исследовать гены, но и произвольно перемещать их из одной клетки в другую, обеспечивает быстрый дальнейший прогресс в иммунологии.
Р.Койко, Д.Саншайн, Э.Бенджамини
Опубликовал Константин Моканов
Источник