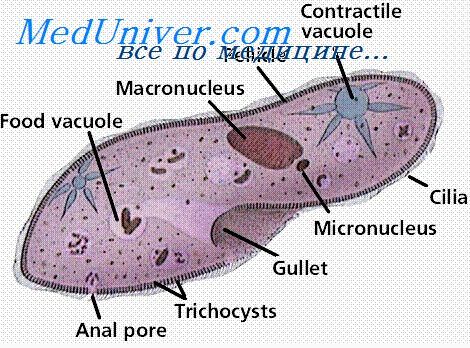
Иммунитет при протозойных инфекциях
Иммуниитет при поражении простейшими. Естественный иммунитет против протозойных инфекцийКак и при всех других инфекциях, при болезнях, вызываемых простейшими (Protozoa), инфекционный процесс и развивающийся по ходу его приобретенный иммунитет формируются на основе взаимоотношений хозяина и паразита. Судьба возбудителя инфекции, проникающего в организм животного или человека, в первую очередь зависит от иммунологического состояния хозяина, которое Ш. Д. Мошковский характеризует как совокупность свойств макроорганизма, определяющих его взаимоотношения с данным возбудителем. Состояние организма хозяина до встречи с антигенами возбудителя (а также с антигенами неживой природы) обозначается Ш. Д. Мошковским как первичное, после встречи — как вторичное иммунологическое. Принципиальное различие между обоими формами иммунологического состояния заключается в том, что при вторичном иммунологическом состоянии к комплексу неспецифических факторов защиты присоединяются факторы специфической защиты — антитела. Абсолютный иммунитет обусловлен полным несоответствием условий в организме хозяина для развития и размножения возбудителей инфекции.
Одним из важнейших факторов, определяющих напряженность относительного иммунитета, является возраст. При ряде протозойных инфекций резистентность организма хозяина с возрастом значительно меняется. Некоторые инфекции у детей и молодых животных протекают тяжело, часто со смертельным исходом, в то время как у взрослых наблюдается более легкое, иногда бессимптомное течение. Примером может служить американский трипаносомоз (болезнь Шагаса). Инфекция Plasmodium berghei, смертельная для молодых крысят, у большинства взрослых крыс протекает доброкачественно. При некоторых инфекциях наблюдаются обратные отношения. Так, например, при бабезиозе (Meitz, 1956) жеребята, ягнята, козлята, поросята до 3-месячного возраста проявляют высокую степень резистентности к соответствующим видам бабезий. Щенята, наоборот, очень чувствительны к этой инфекции. Предполагается, что эти различия обусловлены разной быстротой обратного развития тимуса, которому в последнее время приписывается значительная роль в формировании иммунитета. У щенят атрофия зобной железы наступает через несколько дней после рождения, у других животных — через несколько месяцев. Большая роль в иммунитете к протозойным инфекциям принадлежит селезенке. Skrabolo и Deanovic (1957) описали у человека с удаленной селезенкой тяжелый пироплазмоз со смертельным исходом, вызванный Babesia bovis. Лица, не лишенные этого органа, к пироплазмозу невосприимчивы. Garnham и Вгау (1959) и Вгау и Garnham (1961) наблюдали тяжелую инфекцию Babesia divergans у шимпанзе только после спленэктомни. Хорошо изучена роль селезенки при малярии — экстирпация этого органа у людей, а также у обезьян в латентной (после серии первичных приступов) фазе инфекции приводит к развитию тяжелого, часто смертельного рецидива. Известна различная степень восприимчивости у разных пород животных одного вида к одному и тому же возбудителю: у крупного рогатого скота породы зебу высокая чувствительность к Trypanosoma vivax и значительно меньшая у карликовой породы рогатого скота — ндама. Беспородные белые мыши, а также мыши разных линий высоко чувствительны к Plasmodium berghei, вызывающему у них острую смертельную инфекцию. Но у одной породы мышей — NMRI, наблюдается выраженная резистентность к данному паразиту — инфекция протекает легко и заканчивается выздоровлением (Kretschmar, 1962). Простейшие, как и возбудители других инфекций и инвазий, для своего нормального развития и размножения в организме зараженного хозяина нуждаются в витаминах и других веществах, исключение которых из раниона больного или животного способствует более легкому течению и даже выздоровлению. Taк Maegraith и др., Hawking (1953, 1954) в опытах с Р. bеrghei показали, что у зараженных крыс, содержавшихся исключительно на маточной диете, инфекция не развивается. При добавлении парааминобензойной кислоты инфекция у крыс развивалась нормально. То же установлено и при некоторых формах малярии обезьян — P. knowlesi (Singh и др., 1953), P. cynomolgi (Вгау и Garnham, 1953; Hawking, 1954). — Также рекомендуем «Роль питания в развитии протозойных инфекций. Преодоление иммунитета простейшими» Оглавление темы «Противопротозойный иммунитет. Малярия»: |
Источник
ÐаÑазиÑиÑование пÑоÑÑейÑÐ¸Ñ Ð² оÑганизме Ñеловека и живоÑнÑÑ ÑÑимÑлиÑÑÐµÑ ÑÑнкÑиониÑование гÑмоÑалÑнÑÑ Ð¸ клеÑоÑнÑÑ Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ð¾Ð² иммÑниÑеÑа. Ðднако Ð¸Ñ Ð¿ÑоÑекÑÐ¸Ð²Ð½Ð°Ñ ÑÐ¾Ð»Ñ Ð¿Ñи ÑазлиÑнÑÑ Ð¿ÑоÑозойнÑÑ Ð¸Ð½ÑекÑиÑÑ Ð½ÐµÐ¾Ð´Ð¸Ð½Ð°ÐºÐ¾Ð²Ð°. Ðна в знаÑиÑелÑной меÑе обÑÑловлена ÑизиологиÑеÑкими оÑобенноÑÑÑми ÑÐ°Ð¼Ð¸Ñ Ð¿Ð°ÑазиÑов, Ð¸Ñ Ð¶Ð¸Ð·Ð½ÐµÐ½Ð½Ñм Ñиклом, а Ñакже взаимооÑноÑениÑми Ð¼ÐµÐ¶Ð´Ñ Ð¿Ð°ÑазиÑом и Ñ Ð¾Ð·Ñином, ÑоÑмиÑÑÑÑимиÑÑ Ð² пÑоÑеÑÑе ÑволÑÑионного ÑÑÐ°Ð½Ð¾Ð²Ð»ÐµÐ½Ð¸Ñ Ñой или иной инÑекÑии.
Ðак пÑавило, паÑазиÑиÑование пÑоÑÑейÑÐ¸Ñ Ð¿ÑÐ¸Ð²Ð¾Ð´Ð¸Ñ Ðº поÑÐ²Ð»ÐµÐ½Ð¸Ñ Ð² кÑови позвоноÑного Ñ Ð¾Ð·Ñина ÑпеÑиÑиÑеÑÐºÐ¸Ñ Ð°Ð½ÑиÑел IgM и IgG, коÑоÑÑе обнаÑÑживаÑÑÑÑ Ð¿Ñи помоÑи ÑеÑологиÑеÑÐºÐ¸Ñ ÑеакÑий: ÑвÑзÑÐ²Ð°Ð½Ð¸Ñ ÐºÐ¾Ð¼Ð¿Ð»ÐµÐ¼ÐµÐ½Ñа, иммÑноÑлÑоÑеÑÑенÑии и дÑ.
ÐекоÑоÑÑе пÑоÑÑейÑие (напÑимеÑ, аÑÑиканÑкие ÑÑипаноÑÐ¾Ð¼Ñ — возбÑдиÑели Ñонной болезни) Ñ Ð°ÑакÑеÑизÑÑÑÑÑ Ð²ÑÑокой ÑÑепенÑÑ Ð¸Ð·Ð¼ÐµÐ½ÑивоÑÑи повеÑÑ Ð½Ð¾ÑÑнÑÑ Ð°Ð½Ñигенов, ÑÑимÑлиÑÑÑÑÐ¸Ñ Ð¾Ð±Ñазование ÑзкоÑпеÑиÑиÑеÑÐºÐ¸Ñ Ð°Ð½ÑиÑел, коÑоÑÑе ÑеагиÑÑÑÑ ÑолÑко Ñ Ð¾Ð¿ÑеделеннÑм ваÑианÑом повеÑÑ Ð½Ð¾ÑÑного анÑигена. ÐовÑÑение конÑенÑÑаÑии ÑпеÑиÑиÑеÑÐºÐ¸Ñ Ð°Ð½ÑиÑел Ð²ÐµÐ´ÐµÑ Ðº ÑлиминаÑии ÑÑипаноÑом ÑооÑвеÑÑÑвÑÑÑего анÑигенного ваÑианÑа, на ÑÐ¼ÐµÐ½Ñ ÐºÐ¾ÑоÑÐ¾Ð¼Ñ Ð¿Ð¾ÑвлÑеÑÑÑ Ð½Ð¾Ð²Ñй анÑигеннÑй ваÑÐ¸Ð°Ð½Ñ — и Ñикл повÑоÑÑеÑÑÑ. ÐммÑниÑеÑ, заÑиÑаÑÑий Ð¾Ñ ÐºÐ»Ð¸Ð½Ð¸ÑеÑÐºÐ¸Ñ Ð¿ÑоÑвлений малÑÑии, вÑÑабаÑÑваеÑÑÑ ÑолÑко Ñ Ð»Ð¸Ñ, доÑÑаÑоÑно долго живÑÑÐ¸Ñ Ð² оÑаге болезни и подвеÑгаÑÑÐ¸Ñ ÑÑ Ð¿Ð¾ÑÑоÑнной ÑеинÑекÑии поÑÑедÑÑвом ÑкÑÑов заÑаженнÑÑ Ð¼Ð°Ð»ÑÑийнÑÑ ÐºÐ¾Ð¼Ð°Ñов.
ÐÑи лейÑÐ¼Ð°Ð½Ð¸Ð¾Ð·Ð°Ñ Ð¿Ð¾Ñвление гÑмоÑалÑнÑÑ Ð°Ð½ÑиÑел IgM и IgG ÑвидеÑелÑÑÑвÑÐµÑ Ð¾ пеÑÑиÑÑиÑовании паÑазиÑа в оÑганизме Ñ Ð¾Ð·Ñина в ÑеÑение какого-Ñо пеÑиода вÑемени. ÐÑи ÑÑом анÑиÑела не пÑепÑÑÑÑвÑÑÑ ÑÑпеÑÐ½Ð¾Ð¼Ñ ÑÐ°Ð·Ð¼Ð½Ð¾Ð¶ÐµÐ½Ð¸Ñ Ð¿Ð°ÑазиÑов и не оказÑваÑÑ Ð·Ð°Ð¼ÐµÑного влиÑÐ½Ð¸Ñ Ð½Ð° паÑогенез болезни. РоÑлиÑие Ð¾Ñ Ñонной болезни и малÑÑии лейÑÐ¼Ð°Ð½Ð¸Ð¾Ð·Ñ Ñ Ð°ÑакÑеÑизÑÑÑÑÑ Ð¿Ð¾Ñвлением поÑÑи Ñ Ð²ÑÐµÑ Ð¿ÐµÑеболевÑÐ¸Ñ Ñ Ð¾ÑоÑо вÑÑаженного ÑÑойкого иммÑниÑеÑа. ÐолагаÑÑ, ÑÑо иммÑниÑÐµÑ Ð¿Ñи лейÑÐ¼Ð°Ð½Ð¸Ð¾Ð·Ð°Ñ Ð¸Ð¼ÐµÐµÑ Â«Ð½ÐµÑÑеÑилÑнÑй» Ñ Ð°ÑакÑеÑ, Ñ.е. ÑвÑзан Ñ Ð±ÐµÑÑимпÑомнÑм, лаÑенÑнÑм пеÑÑиÑÑиÑованием паÑазиÑов (иногда пожизненнÑм). РазвиваÑÑийÑÑ Ð¿Ñи лейÑÐ¼Ð°Ð½Ð¸Ð¾Ð·Ð°Ñ Ð¿ÑоÑекÑивнÑй иммÑниÑеÑ, оÑевидно, не ÑвÑзан Ñ Ð³ÑмоÑалÑнÑми анÑиÑелами. Ð ÑÑÐ¸Ñ ÑлÑÑаÑÑ Ð¿ÑевалиÑÑÑÑее знаÑение пÑÐ¸Ð½Ð°Ð´Ð»ÐµÐ¶Ð¸Ñ ÑенÑибилизиÑованнÑм лимÑоÑиÑам. ÐоÑледние, воздейÑÑвÑÑ Ð½Ð° макÑоÑаги, ÑÑимÑлиÑÑÑÑ ÑпоÑобноÑÑÑ ÑÑÐ¸Ñ ÐºÐ»ÐµÑок пÑоÑивоÑÑоÑÑÑ ÑÐ°Ð·Ð¼Ð½Ð¾Ð¶ÐµÐ½Ð¸Ñ Ð² Ð½Ð¸Ñ Ð°Ð¼Ð°ÑÑÐ¸Ð³Ð¾Ñ Ð»ÐµÐ¹Ñманий. ÐÑедполагаÑÑ Ñакже, ÑÑо ÑенÑибилизиÑованнÑе лимÑоÑиÑÑ Ð¼Ð¾Ð³ÑÑ Ð¾ÐºÐ°Ð·ÑваÑÑ ÑиÑоÑокÑиÑеÑкое дейÑÑвие на заÑаженнÑе лейÑманиÑми макÑоÑаги и на паÑазиÑов, оÑвобождаÑÑÐ¸Ñ ÑÑ Ð¿Ñи Ð¸Ñ ÑазÑÑÑении.
ÐкÑивноÑÑÑ ÑÑнкÑиониÑÐ¾Ð²Ð°Ð½Ð¸Ñ Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ð¾Ð² клеÑоÑного иммÑниÑеÑа, Ñ Ð¾Ð´Ð½Ð¾Ð¹ ÑÑоÑонÑ, и ÑоÑÑоÑние иммÑнологиÑеÑкой ÑолеÑанÑноÑÑи — Ñ Ð´ÑÑгой, пÑи лейÑÐ¼Ð°Ð½Ð¸Ð¾Ð·Ð°Ñ , ÑокÑÐ¾Ð¿Ð»Ð°Ð·Ð¼Ð¾Ð·Ð°Ñ Ð¸ дÑÑÐ³Ð¸Ñ Ð¿ÑоÑозойнÑÑ Ð¸Ð½ÑекÑиÑÑ Ð²ÑÑвлÑÑÑ Ð¿Ð¾ налиÑÐ¸Ñ Ð¸Ð»Ð¸ оÑÑÑÑÑÑÐ²Ð¸Ñ ÑеакÑии гипеÑÑÑвÑÑвиÑелÑноÑÑи замедленного Ñипа (ÐÐТ). Так, напÑимеÑ, оÑÑиÑаÑелÑÐ½Ð°Ñ ÑеакÑÐ¸Ñ ÐÐТ, как пÑавило, вÑÑвлÑеÑÑÑ Ñ Ð±Ð¾Ð»ÑнÑÑ Ð¸Ð½Ð´Ð¸Ð¹Ñким кала-азаÑом, ÑÑиопÑким кожнÑм лейÑманиозом и некоÑоÑÑми дÑÑгими ÑоÑмами ÑÑой гÑÑÐ¿Ð¿Ñ Ð·Ð°Ð±Ð¾Ð»ÐµÐ²Ð°Ð½Ð¸Ð¹.
ÐнÑÐ¸Ð³ÐµÐ½Ð½Ð°Ñ Ð¸Ð·Ð¼ÐµÐ½ÑивоÑÑÑ Ð² ÑеÑение жизненного Ñикла, ÑвойÑÑÐ²ÐµÐ½Ð½Ð°Ñ Ð¼Ð½Ð¾Ð³Ð¸Ð¼ паÑазиÑиÑеÑким пÑоÑÑейÑим, а Ñакже пÑевалиÑование клеÑоÑнÑÑ Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ð¾Ð² над гÑмоÑалÑнÑми пÑи ÑоÑмиÑовании пÑоÑекÑивного иммÑниÑеÑа ÑÑезвÑÑайно оÑложнÑÑÑ Ð·Ð°Ð´Ð°ÑÑ ÑÐ¾Ð·Ð´Ð°Ð½Ð¸Ñ ÑÑÑекÑивнÑÑ Ð²Ð°ÐºÑин пÑоÑив пÑоÑозойнÑÑ Ð¸Ð½ÑекÑий.
Также в Ñазделе: ÐÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ñ ÑпеÑиÑиÑеÑкого иммÑниÑеÑа:
Источник
ИММУНИТЕТ ПРИ ПРОТОЗОЙНЫХ БОЛЕЗНЯХ
ÐÐÐУÐÐТÐТ ÐÐ Ð ÐÐ ÐТÐÐÐÐÐЫХ ÐÐÐÐÐÐЯХ
Â
ÐммÑниÑÐµÑ Ðº возбÑдиÑелÑм пÑоÑозойнÑÑ Ð±Ð¾Ð»ÐµÐ·Ð½ÐµÐ¹ Ð¼Ð¾Ð¶ÐµÑ Ð±ÑÑÑ ÐµÑÑеÑÑвеннÑм (вÑожденнÑй) пли пÑиобÑеÑеннÑм.
Â
ÐÑÑеÑÑвеннÑй иммÑниÑÐµÑ Ð²Ð¾Ð·Ð½Ð¸ÐºÐ°ÐµÑ Ð² ÑÐ¸Ð»Ñ Ð±Ð¸Ð¾Ð»Ð¾Ð³Ð¾-ÑкологиÑеÑÐºÐ¸Ñ Ð¾ÑобенноÑÑей оÑганизма живоÑного и ÑвлÑеÑÑÑ Ð²Ð¸Ð´Ð¾Ð²Ñм пÑизнаком Ñ Ð¾Ð·Ñина. ÐÑÑеÑÑвеннÑй иммÑниÑÐµÑ Ðº возбÑдиÑелÑм Ð¼Ð¾Ð¶ÐµÑ Ð±ÑÑÑ Ð°Ð±ÑолÑÑнÑм или оÑноÑиÑелÑнÑм. ÐбÑолÑÑнÑй иммÑниÑÐµÑ Ð½Ð°Ð±Ð»ÑдаÑÑ Ñ ÑÐµÑ Ð¶Ð¸Ð²Ð¾ÑнÑÑ , коÑоÑÑе в лÑбом возÑаÑÑе и пÑи лÑбÑÑ ÑÑловиÑÑ ÑÐ½Ð¸Ð¶ÐµÐ½Ð¸Ñ ÑезиÑÑенÑноÑÑи оÑганизма оÑÑаÑÑÑÑ Ð½ÐµÐ²Ð¾ÑпÑиимÑивÑми к возбÑдиÑелÑм пÑоÑозойнÑÑ Ð±Ð¾Ð»ÐµÐ·Ð½ÐµÐ¹. ÐапÑимеÑ, кÑÑпнÑй ÑогаÑÑй ÑÐºÐ¾Ñ Ð½Ðµ заÑажаеÑÑÑ Ð²Ð¾Ð·Ð±ÑдиÑелем нÑÑÑаллиоза лоÑадей. ÐÑноÑиÑелÑнÑй иммÑниÑÐµÑ Ñ Ð¶Ð¸Ð²Ð¾ÑнÑÑ Ðº возбÑдиÑелÑм пÑоÑозойнÑÑ Ð±Ð¾Ð»ÐµÐ·Ð½ÐµÐ¹ Ñакже возможен. Ð ÑÑом ÑлÑÑае взÑоÑлÑе живоÑнÑе оÑделÑнÑÑ Ð²Ð¸Ð´Ð¾Ð² не воÑпÑиимÑÐ¸Ð²Ñ Ðº возбÑдиÑелÑ, Ñогда как молоднÑк ÑÑого же вида можно заÑазиÑÑ Ð²Ð¾Ð·Ð±ÑдиÑелем. Так, взÑоÑлÑе мÑÑи не воÑпÑиимÑÐ¸Ð²Ñ Ðº возбÑдиÑелÑм пиÑоплазмоза лоÑадей и Ñобак, но новоÑожденнÑе мÑÑи заÑажаÑÑÑÑ Ð¸Ð¼Ð¸.
Â
ÐÑиобÑеÑеннÑй иммÑниÑÐµÑ Ðº возбÑдиÑелÑм пÑоÑозойнÑÑ Ð±Ð¾Ð»ÐµÐ·Ð½ÐµÐ¹ Ñ Ð¶Ð¸Ð²Ð¾ÑнÑÑ Ð²Ð¾Ð·Ð½Ð¸ÐºÐ°ÐµÑ Ð¿Ð¾Ñле пеÑеболеваниÑ, вÑзванного Ñем или дÑÑгим видом паÑазиÑа. РдалÑнейÑем живоÑнÑе ÑÑановÑÑÑÑ Ð½ÐµÐ²Ð¾ÑпÑиимÑивÑми и ÑолÑко к ÑÐ¾Ð¼Ñ Ð²Ð¸Ð´Ñ Ð²Ð¾Ð·Ð±ÑдиÑелÑ, коÑоÑÑй вÑзвал пеÑвиÑное заболевание. ÐÑиобÑеÑеннÑй иммÑниÑÐµÑ Ð¼Ð¾Ð¶ÐµÑ Ð±ÑÑÑ ÑÑеÑилÑнÑм или неÑÑеÑилÑнÑм. СÑеÑилÑнÑй иммÑниÑÐµÑ Ð²Ð¾Ð·Ð½Ð¸ÐºÐ°ÐµÑ Ð² Ñом ÑлÑÑае, когда живоÑное в пÑоÑеÑÑе пеÑÐµÐ±Ð¾Ð»ÐµÐ²Ð°Ð½Ð¸Ñ Ð²ÑÑабаÑÑÐ²Ð°ÐµÑ Ð² оÑганизме анÑиÑела и полноÑÑÑÑ ÑниÑÑÐ¾Ð¶Ð°ÐµÑ Ð²Ð¾Ð·Ð±ÑдиÑÐµÐ»Ñ Ð²Ð¾ вÑÐµÑ Ð¾ÑÐ³Ð°Ð½Ð°Ñ Ð¸ ÑканÑÑ ÑагоÑиÑаÑной акÑивноÑÑÑÑ ÐºÐ»ÐµÑок и дÑÑгими заÑиÑнÑми Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ð°Ð¼Ð¸. ÐеÑÑеÑилÑнÑй иммÑниÑÐµÑ Ð²Ð¾Ð·Ð½Ð¸ÐºÐ°ÐµÑ Ð² Ñом ÑлÑÑае, когда в пÑоÑеÑÑе пеÑÐµÐ±Ð¾Ð»ÐµÐ²Ð°Ð½Ð¸Ñ Ð² оÑганизме живоÑного Ñакже вÑÑабаÑÑваÑÑÑÑ Ð°Ð½ÑиÑела, но ÑагоÑиÑаÑÐ½Ð°Ñ Ð°ÐºÑивноÑÑÑ ÐºÐ»ÐµÑок и дÑÑгие Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ñ Ð·Ð°ÑиÑÑ ÑолÑко подавлÑÑÑ Ð¿Ð°ÑазиÑа, но полноÑÑÑÑ ÐµÐ³Ð¾ не ÑниÑÑожаÑÑ. ÐÑи ÑÑом возбÑдиÑÐµÐ»Ñ Ð´Ð»Ð¸ÑелÑное вÑÐµÐ¼Ñ ÑÐ¾Ñ ÑанÑеÑÑÑ Ð² оÑганизме живоÑного, поддеÑÐ¶Ð¸Ð²Ð°ÐµÑ ÑагоÑиÑаÑнÑÑ Ð°ÐºÑивноÑÑÑ ÐºÐ»ÐµÑок и дÑÑгие заÑиÑнÑе Ð¼ÐµÑ Ð°Ð½Ð¸Ð·Ð¼Ñ Ð¸ Ñем ÑамÑм обÑÑÐ»Ð¾Ð²Ð»Ð¸Ð²Ð°ÐµÑ Ð½ÐµÑÑеÑилÑнÑй иммÑниÑеÑ. ÐÑи пÑоÑозойнÑÑ Ð±Ð¾Ð»ÐµÐ·Ð½ÑÑ ÑаÑе вÑего бÑÐ²Ð°ÐµÑ Ð½ÐµÑÑеÑилÑнÑй иммÑниÑеÑ. ÐлиÑелÑноÑÑÑ ÐµÐ³Ð¾ Ð¾Ñ 4-6 меÑ. до двÑÑ Ð¸ более леÑ. ÐоÑле ÑÑого живоÑное Ð²Ð½Ð¾Ð²Ñ ÑÑановиÑÑÑ Ð²Ð¾ÑпÑиимÑивÑм к Ð´Ð°Ð½Ð½Ð¾Ð¼Ñ Ð²Ð¾Ð·Ð±ÑдиÑелÑ.
← ÐÐÐÐÐÐСТÐÐÐ ÐÐ ÐТÐÐÐÐÐЫХ ÐÐÐÐÐÐÐÐ | ÐÐТÐÐÐÐÐÐ ÐÐ Ð ÐÐ ÐТÐÐÐÐÐЫХ ÐÐÐÐÐÐЯХ → |
|---|
ÐÐ¾Ñ Ð¾Ð¶Ð¸Ð¹ маÑеÑиал по Ñеме:
ÐобавиÑÑ ÐºÐ¾Ð¼Ð¼ÐµÐ½ÑаÑий
Источник