Особенности противовирусного иммунитета микробиология

Ãëàâà 49
Îñîáåííîñòè ïðîòèâîâèðóñíîãî èììóíèòåòà
 îñíîâå íåêîòîðûõ ñóùåñòâåííûõ ðàçëè÷èé ìåõàíèçìîâ çàùèòû îðãàíèçìà îò âèðóñîâ è áàêòåðèé ëåæàò îñîáåííîñòè áèîëîãèè ýòèõ âîçáóäèòåëåé.
 çàùèòå îðãàíèçìà îò âèðóñîâ ó÷àñòâóþò âñå ñèñòåìû èììóíèòåòà, îäíàêî ïðîòèâîâèðóñíûé èììóíèòåò èìååò ñóùåñòâåííûå ñïåöèôè÷åñêèå ÷åðòû. Îíè îïðåäåëÿþòñÿ òåì, ÷òî â ïåðâóþ î÷åðåäü íà ïðîíèêíîâåíèå âèðóñà â îðãàíèçì ðåàãèðóþò íå ñèñòåìû êîìïëåìåíòà è ìàêðîôàãîâ, à ñèñòåìû èíòåðôåðîíîâ è Ò-êèëëåðíûõ êëåòîê. Äðóãàÿ îñîáåííîñòü ôîðìèðîâàíèÿ èììóíèòåòà ñâÿçàíà ñ òåì, ÷òî âèðóñû îêàçûâàþò ñëàáîå àíòèãåííîå âîçäåéñòâèå íà Â-ëèìôîöèòû è äëÿ èõ àêòèâèðîâàíèÿ, ïðîëèôåðàöèè è äèôôåðåíöèðîâêè íåîáõîäèìî ó÷àñòèå Ò-õåëïåðîâ è ñîîòâåòñòâåííî ïðåäñòàâëåíèå ïîñëåäíèì ïðîöåññèðîâàííîãî âèðóñíîãî àíòèãåíà (ïåïòèäíûõ ôðàãìåíòîâ) ïðè ó÷àñòèè ìîëåêóë ÌÍÑ êëàññà II. Ïîýòîìó ðîëü ìàêðîôàãîâ è äðóãèõ àíòèãåíïðåäñòàâëÿþùèõ êëåòîê çàêëþ÷àåòñÿ íå ñòîëüêî â ñàìîì ôàãîöèòîçå, ñêîëüêî â ïðîöåññèðîâàíèè è ïðåäñòàâëåíèè àíòèãåíà.
Åùå îäíà îñîáåííîñòü ïðîòèâîâèðóñíîãî èììóíèòåòà îáóñëîâëåíà ñòðóêòóðíîé îðãàíèçàöèåé íåêîòîðûõ âèðèîíîâ. Âèðóñû ìîãóò âûçâàòü çàáîëåíèå ëèøü â òîì ñëó÷àå, åñëè ïðîíèêàþò â êëåòêó. Äëÿ ïðèêðåïëåíèÿ ê íåé îíè èñïîëüçóþò êëåòî÷íûå ðåöåïòîðû, êîòîðûå êëåòêà èñïîëüçóåò äëÿ ñîáñòâåííûõ ôèçèîëîãè÷åñêèõ öåëåé. Âñå èäåíòèôèöèðîâàííûå âèðóññïåöèôè÷åñêèå ðåöåïòîðû ãëèêîïðîòåèäû èëè ñèàëîãëèêîëèïèäû. Âèðóñ «óçíàåò» ñïåöèôè÷åñêèå ðåöåïòîðû è ïðèêðåïëÿåòñÿ ê íèì ñ ïîìîùüþ ñâîèõ ïðèêðåïèòåëüíûõ áåëêîâ VAP (àíãë. virion attachment proteins). Èìåííî îíè èãðàþò ðîëü ñâîåîáðàçíûõ ëîöìàíîâ, íàïðàâëÿþùèõ äâèæåíèå âèðóñà â êëåòêó. Ó íåêîòîðûõ âèðóñîâ ìîëåêóëû ýòèõ áåëêîâ-ëîöìàíîâ ðàñïîëîæåíû â ñêðûòûõ ìåñòàõ «ùåëÿõ», «êàíüîíàõ», ò. å. óãëóáëåíèÿõ íà ïîâåðõíîñòè âèðèîíà. Èõ äèàìåòð (ãëóáèíà) ó âèðóñîâ ãðèïïà, ïîëèîìèåëèòà, ÂÈ× íå ïðåâûøàåò 2,5 íì. Äèàìåòð æå àêòèâíîãî öåíòðà ìîëåêóëû àíòèòåëà ñîñòàâëÿåò 3,5 íì, ïîýòîìó àíòèòåëî íå ìîæåò ñâÿçàòüñÿ ñ áåëêîì-ëîöìàíîì âèðóñà è áëîêèðîâàòü åãî.  ðåçóëüòàòå âèðóñíåéòðàëèçóþùàÿ àêòèâíîñòü àíòèòåë îñëàáëÿåòñÿ. Íà ïðîíèêíîâåíèå âèðóñà ðàíüøå âñåãî ðåàãèðóåò ñèñòåìà èíòåðôåðîíîâ, êîòîðûå ïîäàâëÿþò âíóòðèêëåòî÷íîå ðàçìíîæåíèå âèðóñîâ. Êðîìå òîãî, ïðîòèâîâèðóñíîå äåéñòâèå îêàçûâàþò íàõîäÿùèåñÿ â ñûâîðîòêå êðîâè ? è ?-èíãèáèòîðû. Àëüôà-èíãèáèòîð òåðìîñòàáèëüíûé ñóáñòðàò, âõîäèò â ñîñòàâ ?-ãëîáóëèíîâ, ïðåïÿòñòâóåò àäñîðáöèè âèðóñîâ íà êëåòêå, ðàçðóøàåòñÿ íåéðàìèíèäàçîé îðòî è ïàðàìèêñîâèðóñîâ. Áåòà-èíãèáèòîð òåðìîëàáèëüíûé ìóêîïåïòèä, âõîäèò â ñîñòàâ ?-ãëîáóëèíîâ, ïîäàâëÿåò ðàçìíîæåíèå îðòî è ïàðàìèêñîâèðóñîâ.
Îäíàêî èíòåðôåðîíîâ è èíãèáèòîðîâ îêàçàëîñü íåäîñòàòî÷íî äëÿ çàùèòû îò âèðóñîâ, ïîýòîìó ïðèðîäà ñîçäàëà ïðîòèâ âèðóñîâ äðóãîé, î÷åíü ìîùíûé ìåõàíèçì çàùèòû íà óðîâíå îðãàíèçìà. Îí ïðåäñòàâëåí ïðåæäå âñåãî Ò-öèòîòîêñè÷åñêèìè ëèìôîöèòàìè è äðóãèìè êèëëåðíûìè êëåòêàìè. Ýòè êëåòêè ðàñïîçíàþò âñå ÷óæåðîäíûå àíòèãåíû, â òîì ÷èñëå è âèðóñíûå, ïðåäñòàëÿåìûå èì ìîëåêóëàìè ÌÍÑ êëàññà I. Ãëàâíîå áèîëîãè÷åñêîå çíà÷åíèå Ò-êèëëåðíûõ êëåòîê è çàêëþ÷àåòñÿ â îáíàðóæåíèè è óíè÷òîæåíèè ëþáûõ êëåòîê, èíôèöèðîâàííûõ ÷óæåðîäíûìè àíòèãåíàìè.
Ñèíòåç àíòèòåë ñâÿçàí, â ñâîþ î÷åðåäü, ñ ñèñòåìîé ôàãîöèòîâ,  è Ò-ëèìôîöèòîâ è ÌÍÑ. Â-ëèìôîöèòû ñ ïîìîùüþ èììóíîãëîáóëèíîâûõ ðåöåïòîðîâ ðàñïîçíàþò ñîîòâåòñòâóþùèé àíòèãåí è îòâå÷àþò íà íåãî ñèíòåçîì ðåöåïòîðîâ, íåîáõîäèìûõ äëÿ ðàñïîçíàâàíèÿ ñèãíàëîâ îò Ò-õåëïåðîâ. Àêòèâèðîâàííûå Ò-õåëïåðû ñèíòåçèðóþò è ñåêðåòèðóþò ôàêòîðû àêòèâàöèè, ïðîëèôåðàöèè è äèôôåðåíöèàöèè Â-ëèìôîöèòîâ.  ðåçóëüòàòå èõ äåéñòâèÿ èç àêòèâèðîâàííûõ Â-ëèìôîöèòîâ ôîðìèðóþòñÿ êëîíû àíòèòåëîîáðàçóþùèõ êëåòîê è êëåòîê ïàìÿòè (ñîîòâåòñòâåííî âîçíèêàþò è êëîíû êëåòîê ïàìÿòè Ò-ëèìôîöèòîâ).
Çàùèòíàÿ ðîëü àíòèòåë â ïðîòèâîâèðóñíîì èììóíèòåòå ñîñòîèò ãëàâíûì îáðàçîì â òîì, ÷òî îíè, âçàèìîäåéñòâóÿ ñ âèðóñíûìè ðåöåïòîðàìè, èñêëþ÷àþò âîçìîæíîñòü àäñîðáöèè âèðóñîâ íà ìåìáðàíå êëåòîê è òàêèì îáðàçîì íåéòðàëèçóþò èõ àêòèâíîñòü, äåëàþò íåâîçìîæíûì ïðîíèêíîâåíèå âèðóñà â êëåòêó. Òîëüêî òàêèì ïóòåì, ò. å. èñêëþ÷àÿ âîçìîæíîñòü ïðîíèêíîâåíèÿ âèðóñà â êëåòêó, àíòèòåëà îáåñïå÷èâàþò ôîðìèðîâàíèå ïðèîáðåòåííîãî èììóíèòåòà. Ïðèîáðåòåííûé ïðîòèâîâèðóñíûé èììóíèòåò ïðè íàëè÷èè ïîñòîÿííûõ êëîíîâ êëåòîê ïàìÿòè ìîæåò ñîõðàíÿòüñÿ ïîæèçíåííî. Ïîìèìî ñïîñîáíîñòè íåéòðàëèçîâàòü âèðóñû, àíòèòåëà âûïîëíÿþò áîëüøóþ ðîëü â îñâîáîæäåíèè îðãàíèçìà îò âèðóñîâ è âèðóñíûõ àíòèãåíîâ. Ñâÿçûâàÿñü ñ íèìè, àíòèòåëà îáðàçóþò èììóííûå êîìïëåêñû, êîòîðûå âûâîäÿò ýòè àíòèãåíû èç îðãàíèçìà. Ýôôåêòèâíîñòü àíòèòåë â ôîðìèðîâàíèè ïðèîáðåòåííîãî èììóíèòåòà ïðîòèâ âèðóñíûõ èíôåêöèé ïîäòâåðæäåíà ìíîãîëåòíåé ïðàêòèêîé ñïåöèôè÷åñêîé ïðîôèëàêòèêè ïîëèîìèåëèòà, êîðè, æåëòîé ëèõîðàäêè, äðóãèõ èíôåêöèé è ïîëíîé ëèêâèäàöèåé íà Çåìëå íàòóðàëüíîé îñïû.
Âìåñòå ñ òåì ñàìè ïî ñåáå âèðóñû ìîãóò ñòàòü ïðè÷èíîé âòîðè÷íûõ èììóíîäåôèöèòîâ. Òèïè÷íûì ïðèìåðîì òàêîãî âèðóñíîãî èììóíîäåôèöèòà ÿâëÿåòñÿ ÂÈ×-èíôåêöèÿ (ñì. ãëàâó 57).
Источник
Особенности противовирусного иммунитета обусловлены своеобразием анатомического строения вирусов, сравнительно небольшим набором антигенов их оболочек, возможностью дрейфа поверхностных антигенов (белков), абсолютным паразитизмом вирусов, особенностью их взаимодействия с чувствительными клетками.
В макроорганизме вирус может находиться в различных состояниях:
a) внеклеточно (вирион);
b) внутриклеточно, на разных стадиях быстрого или медленного продуктивного взаимодействия с чувствительной клеткой (вирус);
c) быть интегрированным в геном клетки-мишени (непродуктивное взаимодействие, провирус).
Соответственно этим основным состояниям вируса формирующийся противовирусный иммунитет направлен на нейтрализацию и удаление вируса и его антигенов из организма, что достигается с помощью антител, а также на уничтожение собственных инфицированных вирусом клеток-мишеней цитотоксическими Т-лимфоцитами (Тс).
Образующиеся при вирусных инфекциях антитела IgG могут участвовать в разных биологических реакциях.
1. Нейтрализация инвазивных свойств вирионов.Образовавшийся комплекс связывается с поверхностью макрофага за счет его Fc-рецепторов. Поглощение комплекса обычно ведет к гибели возбудителя (рис. 58), непоглощенные иммунные комплексы могут диссоциировать, а освободившиеся вирионы заражают чувствительные клетки. Длительная циркуляция непоглощенных и недиссоциированных иммунных комплексов по всему организму может приводить к депонированию их в различных тканях организма и индуцировать развитие местных воспалительных реакций через активацию системы комплемента или интерлейкинов после фиксации комплекса клетками, имеющими рецептор к Fc-фрагменту антител (гепатит В, инфекционный мононуклеоз, подострый склерозирующий панэнцефалит и др.).
2. Антителоопосредованный комплемент-зависимый цитолиз зараженных вирусом клеток-мишеней представлен на рис. 58. Лизис мембраны зараженной клетки происходит за счет мембраноатакующего комплекса (МАК) комплемента. Освободившиеся вирионы подвергаются воздействию антител.
3. Антителоопосредованный цитолизклеток-мишеней макрофагами и гранулоцитами при выделении ими в момент контакта с пораженной клеткой гранзимов и цитолизинов (рис. 58). Такие макрофаги и гранулоциты должны иметь Fc-рецепторы. Специфичностью по отношению к вирусному антигену они не обладают. Цитотоксические Т-лимфоциты в этой реакции не участвуют. Их активность от наличия антител не зависит. Разрушение пораженных вирусом клеток осуществляется также цитотоксическими Т-лимфоцитами Тс (рис. 59). Тс способны лизировать инфицированные вирусом клетки, реагируя на вирусный антиген представленный клеткой на ГКГС-I.
Рис. 58. Участие АТ в противовирусном иммунитете
а) Нейтрализация антителами свободных вирионов.
б) Антителопосредованный комплиментзависимый цитоз клетки – мишени.
в) Антителопосредованный цитоз гранулоцитами.
Для цитотоксического действия Тс-лимфоцитам необходим непосредственный контакт с клеткой-мишенью. После этого происходит выделение Тс-лимфоцитом гранзимов или цитолизинов, вызывающих изменение мембранной проницаемости клетки-мишени. Ее осмотическое набухание, разрыв мембраны и выход содержимого цитоплазмы в микроокружающую среду.
Рис. 59. Взаимодействие цитотоксического
Т-лимфоцита с клеткой – мишенью.
Способностью к интеграции вирусной нуклеиновой кислоты в геном клетки-мишени обладают ДНК-содержащие вирусы и ретровирусы. Потомство зараженной клетки наследует провирус. Вирусные антигены (белки) в клетке не синтезируются, они не представлены на ГКГС-I. Иммунному надзору такая клетка не поддается.
ОСНОВНАЯ ЛИТЕРАТУРА
1. Авакян А.А., Быковский А.Ф. Атлас анатомии и онтогенеза вирусов человека и животных. М., 1970.
2. Бережная Н.М., Бобкова Л.П., Петровская И.А., Ялкут С.И. Аллергология. – Киев. – 1986. – 445с.
3. Вирусология. Под ред. Б. Филдса и Д. Найпа. – т. 1. – М. «Мир», 1989. – 494с.
4. Генис Д.В. Медицинская паразитология. М., Медицина. – 1991. – 240с.
5. Гусев М.В., Минеева Л.А. Микробиология. М., МГУ. – 1994. – 294с.
6. Джавец Э., Мелькик Дж. Л., Эйдельберт Э.А. Руководство по медицинской микробиологии. М., 1982, Т.1. – 264с.
7. Западнюк И.П., Западнюк В.И., Захарня Е.А. Лабораторные животные. – Киев. – 1983. – 292с.
8. Кашкин П.Н., Лисин В.В. Практическое руководство по медицинской микологии. М., 1983. – 185с.
9. Коротяев А.И., Бабичев С.А. Медицинская микробиология с иммунологией и вирусологией. С.-Петербург, 1998. -–592с.
10. Льюин Б. Гены. – Мир, 1987. – 544с.
11. Маянский А.Н. Микробиология для врачей. Издательство НГМА, Нижний Новгород, 1999. – 450с.
12. Медицинская микробиология. Под ред. В.И. Покровского и О.Е. Поздеева. М.- «Геотар медицина» — 1998. – 1184с.
13. Микробиология и иммунология. Под ред. А.А. Воробьева. М. – «Медицина» — 1999. – 464с.
14. Плейфер Дж. Наглядная иммунология. М. – ГЭОТАР Медицина. – 1999. – 96с.
15. Шлегель Г. Общая микробиология. – М., Мир. – 1987. – 566с.
Источник
Особенности
противовирусного иммунитета обусловлены
своеобразием анатомического строения
вирусов, сравнительно небольшим набором
антигенов их оболочек, возможностью
дрейфа поверхностных антигенов (белков),
абсолютным паразитизмом вирусов,
особенностью их взаимодействия с
чувствительными клетками.
В
макроорганизме вирус может находиться
в различных состояниях:
внеклеточно
(вирион);внутриклеточно,
на разных стадиях быстрого или медленного
продуктивного взаимодействия с
чувствительной клеткой (вирус);быть интегрированным
в геном клетки-мишени (непродуктивное
взаимодействие, провирус).
Соответственно
этим основным состояниям вируса
формирующийся противовирусный иммунитет
направлен на нейтрализацию и удаление
вируса и его антигенов из организма,
что достигается с помощью антител, а
также на уничтожение собственных
инфицированных вирусом клеток-мишеней
цитотоксическими Т-лимфоцитами (Тс).
Образующиеся
при вирусных инфекциях антитела IgGмогут участвовать в разных биологических
реакциях.
1.
Нейтрализация инвазивных свойств
вирионов. Образовавшийся комплекс
связывается с поверхностью макрофага
за счет егоFc-рецепторов.
Поглощение комплекса обычно ведет к
гибели возбудителя (рис. 58), непоглощенные
иммунные комплексы могут диссоциировать,
а освободившиеся вирионы заражают
чувствительные клетки. Длительная
циркуляция непоглощенных и недиссоциированных
иммунных комплексов по всему организму
может приводить к депонированию их в
различных тканях организма и индуцировать
развитие местных воспалительных реакций
через активацию системы комплемента
или интерлейкинов после фиксации
комплекса клетками, имеющими рецептор
кFc-фрагменту антител
(гепатит В, инфекционный мононуклеоз,
подострый склерозирующий панэнцефалит
и др.).
2.
Антителоопосредованный комплемент-зависимый
цитолиззараженных вирусом клеток-мишеней
представлен на рис. 58. Лизис мембраны
зараженной клетки происходит за счет
мембраноатакующего комплекса (МАК)
комплемента. Освободившиеся вирионы
подвергаются воздействию антител.
3.
Антителоопосредованный цитолиз
клеток-мишеней макрофагами и
гранулоцитами при выделении ими в момент
контакта с пораженной клеткой гранзимов
и цитолизинов (рис. 58). Такие макрофаги
и гранулоциты должны иметьFc-рецепторы.
Специфичностью по отношению к вирусному
антигену они не обладают. Цитотоксические
Т-лимфоциты в этой реакции не участвуют.
Их активность от наличия антител не
зависит. Разрушение пораженных вирусом
клеток осуществляется также цитотоксическими
Т-лимфоцитами Тс (рис. 59). Тс способны
лизировать инфицированные вирусом
клетки, реагируя на вирусный антиген
представленный клеткой на ГКГС-I.
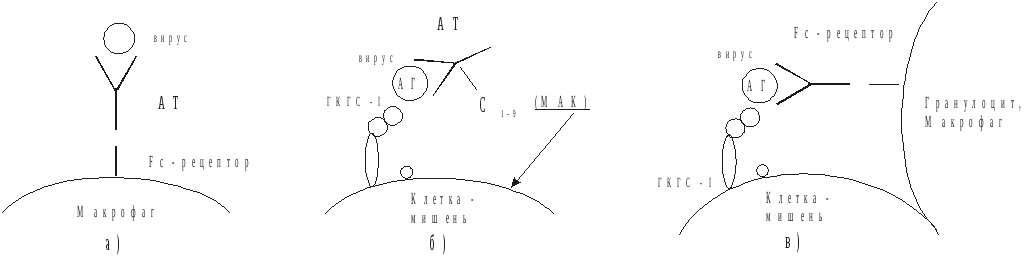
Рис. 58. Участие АТ в противовирусном
иммунитете
а) Нейтрализация антителами свободных
вирионов.
б) Антителопосредованный комплиментзависимый
цитоз клетки – мишени.
в) Антителопосредованный цитоз
гранулоцитами.
Д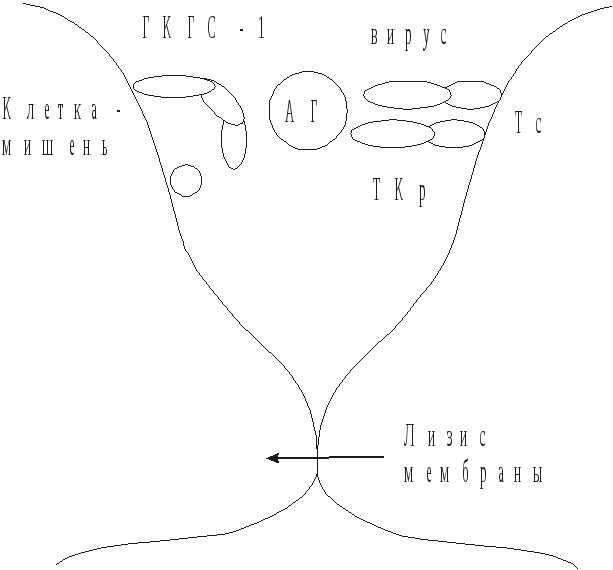 ля
ля
цитотоксического действия Тс-лимфоцитам
необходим непосредственный контакт с
клеткой-мишенью. После этого происходит
выделение Тс-лимфоцитом гранзимов или
цитолизинов, вызывающих изменение
мембранной проницаемости клетки-мишени.
Ее осмотическое набухание, разрыв
мембраны и выход содержимого цитоплазмы
в микроокружающую среду.
Рис. 59.
Взаимодействие цитотоксического
Т-лимфоцита
с клеткой – мишенью.
Способностью
к интеграции вирусной нуклеиновой
кислоты в геном клетки-мишени обладают
ДНК-содержащие вирусы и ретровирусы.
Потомство зараженной клетки наследует
провирус. Вирусные антигены (белки) в
клетке не синтезируются, они не
представлены на ГКГС-I.
Иммунному надзору такая клетка не
поддается.
ОСНОВНАЯ ЛИТЕРАТУРА
Авакян А.А., Быковский
А.Ф. Атлас анатомии и онтогенеза вирусов
человека и животных. М., 1970.Бережная Н.М.,
Бобкова Л.П., Петровская И.А., Ялкут С.И.
Аллергология. – Киев. – 1986. – 445с.Вирусология. Под
ред. Б. Филдса и Д. Найпа. – т. 1. – М.
«Мир», 1989. – 494с.Генис Д.В. Медицинская
паразитология. М., Медицина. – 1991. –
240с.Гусев М.В., Минеева
Л.А. Микробиология. М., МГУ. – 1994. – 294с.Джавец Э., Мелькик
Дж. Л., Эйдельберт Э.А. Руководство по
медицинской микробиологии. М., 1982, Т.1.
– 264с.Западнюк И.П.,
Западнюк В.И., Захарня Е.А. Лабораторные
животные. – Киев. – 1983. – 292с.Кашкин П.Н., Лисин
В.В. Практическое руководство по
медицинской микологии. М., 1983. – 185с.Коротяев А.И.,
Бабичев С.А. Медицинская микробиология
с иммунологией и вирусологией.
С.-Петербург, 1998. -–592с.Льюин Б. Гены. –
Мир, 1987. – 544с.Маянский А.Н.
Микробиология для врачей. Издательство
НГМА, Нижний Новгород, 1999. – 450с.Медицинская
микробиология. Под ред. В.И. Покровского
и О.Е. Поздеева. М.- «Геотар медицина» —
1998. – 1184с.Микробиология и
иммунология. Под ред. А.А. Воробьева. М.
– «Медицина» — 1999. – 464с.Плейфер Дж.
Наглядная иммунология. М. – ГЭОТАР
Медицина. – 1999. – 96с.Шлегель Г. Общая
микробиология. – М., Мир. – 1987. – 566с.
Соседние файлы в предмете Микробиология
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Глава 16. Противовирусный иммунитет
■ Вирусы — эгооблигатныевнутриклеточныепаразиты, чрезвычайно разнообразные по строению и способам размножения. Одни из них вызывают острые вирусные инфекции и в конечном итоге уничтожаются организмом-хозяином, другие неопределенно долго персистируют в нем, вызывая «медленные вирусные инфекции».
■ Факторыврожденногоиммунитета, в частности интерфероны и НК-клетки, препятствуют инфекции на ее ранних стадиях и задерживают распространение вируса по организму.
■ Антителанейтрализуютинфекционностьвирусов, препятствуя их проникновению в новые клетки и ткани, и служат важным механизмом защиты от повторной инфекции.
■ ЦитотоксическиеТ—лимфоциты способны распознавать инфицированные вирусами клетки организма и разрушать их в период ранней стадии репродукции вируса, прежде чем появится новоепоколение вирусных частиц.
■ Вирусыспособныизбегатьраспознавания иммунной системой организма, так как обладают свойствами латентной персистенции, антигенной изменчивости и синтеза «белков-ловушек», препятствующих действию защитных механизмов хозяина.
■ Вирусымогутнепосредственнонарушатьфункциииммуннойсистемы, вызывая иммуносупрессию, иммунологическую недостаточность и аутоиммунные заболевания.
ТИПЫ ВИРУСНЫХ ИНФЕКЦИЙ
Вирусы — ото облигатные внутриклеточные паразиты, использующие для синтеза своих белков биохимический аппарат клетки-хозяина. Они чрезвычайно разнообразны по строению и организации генома — некоторые имеют РНК-геном, состоящий всего из нескольких генов, другие обладают ДНК-геномами с числом генов до двух сотен. Структурно вирус представляет собой просто белковый «футляр» (капсид), в которым упакована нуклеиновая кислота. Обнаружены и еще более простые формы живых организмов:
• вироилы — возбудители болезней растений, состоящие только из нуклеиновой кислоты без белковой оболочки, и
• прионы — «инфекционные белки», ассоциированные с дегенеративными неврологическими болезнями животных и человека, включая скрейпи (почесуху овец), спонгиозные (губчатые) энцефалопатии крупного рогатого скота и болезнь Крейцфельдта Якоба.
Этапы развития типичной вирусной инфекции на уровне клетки показаны на рис. 16.1. Вирусы прикрепляются к клеткам хозяина, связываясь со специфическими клеточными рецепторами. Этой специфичностью обусловлен тропизм данного вируса к определенному виду-хозяину или типу клеток. Примеры клеточных рецепторов, которыми пользуются вирусы, приведены на рис. 16.2. После проникновения вируса в клетку происходит его раздевание дезагрегация белкового капсида с высвобождением нуклеиновой кислоты. Затем начинается транскрипция и синтез вирусных белков, репликация вирусного генома и, наконец, сборка и созревание нового «поколения» вирусных частиц. Выйдя в межклеточное пространство, они инфицируют близлежащие клетки и ткани. В зависимости от вида вируса и особенностей метаболизма клетки-хозяина этот процесс может различаться в деталях. Так, пикорнавирусам (мелкие РНК-вирусы) дня образования новых вирусных частиц требуется около 8 ч, а ДНК-содержащему цитомегаловирусу человека — до 48 ч.

Рис. 16.1. Для размножения вирусу необходимо проникнуть в клетку организма-хозяина.
Рис. 16.2. Вирусы прикрепляются к клеткам, пользуясь специфическими клеточными рецепторами. Специфичностью рецепторов отчасти предопределено, какие типы клеток будут инфицированы.
Механизмы заражения хозяина, персистениии в тканях и патогенеза инфекции широко вирьируют среди различных вирусов. Как правило, вирусы проникают во внутреннюю среду организма через слизистые оболочки. Другой, очень эффективный путь их прямого проникновения в кровоток — через поврежденную кожу, например при укусе насекомого или уколе инъекционной! иглой. Размножение вирусов обычно происходит в эпителиальных покровах; вслед за этим в некоторых случаях вирусы проникают в кровь (виремия), что приводит к их распространению и инфицированию других тканей. Выздоровление может означать полное устранение вируса из организма-хозяина. Однако некоторые вирусы (в частности, вирус герпеса) способны оставаться (персистировать) в организме в скрытой (латентной, неинфекционной) форме после затухания острой инфекции и в какой-то момент реактивироваться с образованием новых инфекционных вирусных частиц. Другие вирусы (например, Rupyc гепатита В или вирус лимфоцитарного хориоменингита) персистируют в инфекционной форме, несмотря на иммунный ответ хозяина. При таких заболеваниях, как скрейпи и болезнь Крейцфельдта-Якоба, вовсе не бывает острой стадии; их возбудители персистируют в организме, что сопровождается медленной инфекцией, и вызывают проявления болезни спустя многие годы после заражения. В отличие от истинных вирусных, прионовые инфекции не стимулируют ни иммунного ответа, ни продукции интерферонов. Классификация вирусных инфекций по клиническому течению представлена на рис. 16.3.

Рис. 16.3. Вирусные инфекции могут протекать остро или хронически и иметь различный исход.
Источник