Роль иммунитета в эволюции

Исходя из тех представлений, что величина мутационного риска прямо пропорциональна количеству делящихся клеток, следует допустить, что параллельно процессу развития многоклеточности шло формирование механизмов, сдерживающих мутационный поток. В данном случае необходимо обратить внимание не на внешние биоценотические факторы, а на внутренние, свойственные самому многоклеточному организму. Без успешного формирования этих механизмов эволюция “застряла” бы на том уровне, при котором количество соматических клеток у какого-либо животного ничтожно мало, а период воспроизведения короток. В этом смысле критическая точка — 106 клеток — гипнотизирует. Она выглядит некоторым пределом в эволюции многоклеточных по линии увеличения количества активно делящихся клеточных форм.
Суммируя представленный в данной главе материал, можно попытаться привести в соответствие факты по эволюционному возникновению различных способов иммунного реагирования с уровнем организации в мире животных (рис. 20.14). Всего включено пять проявлений иммунитета: фагоцитоз, аллогенная ингиби- ция, специфическая клеточная форма защиты, реактивность ан- тигенраспознающих рецепторов и продукция иммуноглобулинов.
Способность одноклеточных организмов к фагоцитозу является тем свойством, которое обеспечивает их питание. Фагоцитоз как реакция амебоцитов-макрофагов на чужеродный материал сохранился у всех многоклеточных животных.
Данные по неиммунному распознаванию чужеродносги у большинства изученных представителей наиболее просто организованных многоклеточных — губок и кишечнополостных (табл. 20.6), а также демонстрация явления аллогенного подавления у млекопитающих (гл. 11) позволяют думать, что подобная форма реактивности есть общее свойство всех многоклеточных.
Факты зарождения клеточных форм специфического реагирования у простейших многоклеточных и усиление специфического клеточного иммунитета , обусловленного активностью лимфоцитов, у первично- и вторичноротых животных определяют рамки, в которых данная форма иммунитета имеет место. Если верхний предел специфического клеточного реагирования ясен (это уровень млекопитающих), то нижний (губки, кишечнополостные) выглядит достаточно расплывчатым. Лишь некоторые представи
ло
Рис 20.!4- Соотношение между различными факторами иммунитета и уровнями организации и мире животных
тели этих таксонов способны к специфическому реагированию с формированием кратковременной иммунологической памяти. Подобные отношения следует отнести к явлению преадаптации элементов клеточной формы реагирования. На рисунке квазииммун- ная форма реагирования отмечена пунктирной линией.
Специфичность клеточной иммунной реакции подразумевает наличие на поверхности эффекторных клеток молекулярных структур, способных к распознаванию чужеродного антигенного материала. Несмотря на то что наличие антигенраспознающих рецепторов у беспозвоночных выявлено только у иглокожих и оболочников, следует предполагать присутствие этих рецепторов также у более низкоорганизованных беспозвоночных, способных к специфическому клеточному реагированию. Широкое распространение оцнодоменных белков суперсемейства иммуноглобулинов (Thy-1, р2-микроглобулина, Р0 — гл. 5) от одноклеточных до высших позвоночных животных вселяет уверенность в том, что какие- то иммуноглобулинподобные, антигенраспознающие структуры будут найдены у низкоорганизованных многоклеточных, способных к специфическому реагированию.
И, наконец, показано, что синтез иммуноглобулинов — компонентов специфического гуморального реагирования — есть привилегия позвоночных животных.
Анализ схемы, представленной на рис. 20.13, ясно указывает на необходимость совершенствования иммунных механизмов кон-
441
троля за процессом эволюционного развития многоклеточных животных.
Конечно, тезис о том, что прогресс по линии увеличения количества соматических клеток обеспечивался системой иммунологического контроля за мутационным потоком, бьы бы более убедительным, если бы удалось провести строгую коррелятивную связь между эволюционно возникающими формами иммунитета и все увеличивающимися размерами носителей этого иммунитета, как это сделано для развивающегося зародыша человека (гл. 19). Отсутствие сведений об истинном количестве воспроизводящихся клеток у представителей разных таксонов, недостаток знаний о состоянии иммунной реактивности у наиболее просто организованных животных не позволяют установить абсолютной связи. И тем не менее, можно попытаться провести подобную оценку в гипотетической форме, опираясь на возможные в прошлом пути развития многоклеточности. На рис. 20.15 в условном графическом виде представлены варианты эволюционных изменений количества возобновляющихся клеток у представителей тех или иных таксономических групп животных. Относительная клеточная величина отображена размером круга (чем больше предполагаемое количество воспроизводящихся клеток, тем больше диаметр круга и наоборот). Условно нарастание многоклеточности обозначено последовательными цифрами I — 9. Следует еще раз подчеркнуть, что данная схема полностью умозрительна и указывает лишь на возможные тенденции в эволюционном развитии многоклеточности.
В глубоком геологическом прошлом (очевидно, в архее) эволюция по линии увеличения количества соматических клеток, вероятно, завершилась бы на стадии С4 и не имела бы успеха в дальнейшем увеличении клеточности по причине отсутствия специфического контроля за мутационным потоком. Вместе с тем, при том же уровне многоклеточности форма D4, обладающая определенным видом специфического иммунологического контроля, “обречена”на эволюционный успех. Дальнейшее историческое развитие могло привести как к увеличению абсолютного количества пролиферирующих клеток (линия D-H), так и к различного рода колебаниям по многоклеточности (D-Dn, Е-Еп и т.д,). Это может быть связано с различными условиями существования вида и действием факторов отбора, отличных от иммунных. В результате на уровне современных форм (Dn-Hn) диапазон колебаний многоклеточности велик, а представители с незначительным количеством клеток, но прошедшие предковый путь становления иммунитета, соседствуют с формами, имеющими большее количество соматических клеток, но не обладающими специфическим иммунитетом. Так, наиболее мелкие виды высокоорганизованных клаЬ- 442
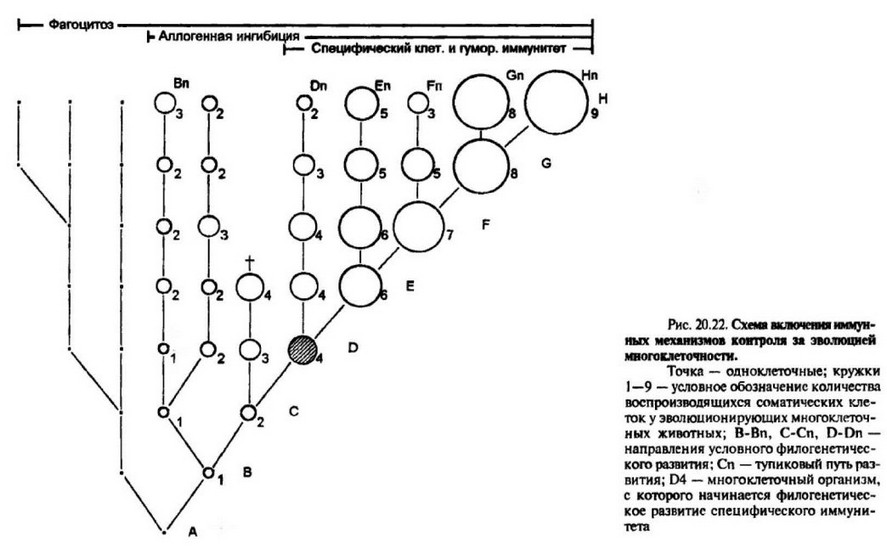
сов птиц и млекопитающих, имеющих полностью сформированные механизмы иммунной защиты, уступают по размерам некоторым видам кишечнополостных, либо не обладающим специфической защитой, либо с очень слабой защитой. Иными словами, выдвигая тезис о контролирующей роли исторически развивающихся механизмов специфического иммунитета, следует относить эту роль к начальным этапам становления многоклеточное™, к тому исходному периоду, когда определялся сам путь эволюционного развития многоклеточное™.
Каково количество соматических клеток, начиная с которого оно не могло бы увеличиваться без параллельного становления специфических форм иммунологической защиты? Исходя из принципов биогенетического закона Геккеля, следует думать, что такое количество незначительно. Уже на самых ранних этапах зародышевого развития позвоночных животных регистрируются признаки Т- и В-систем иммунитета (гл. 19). Возможно, эта величина составляла lxlO6 воспроизводящихся клеток — тог предел, с которого мутационный риск становится положительной величиной (рис. 20.13).
На основании всего сказанного выше следует заключить, что на эволюцию иммунитета не следует смотреть только как на самостоятельную линию исторического развития, связанную с анти- инфекционной защитой, но скорее как на такой эволюционный процесс, который самым тесным образом связан с эволюцией многоклеточных вообще. В связи с подобными представлениями кажется разумным утверждение, согласно которому исторически развивающийся иммунитет явился одним из важных факторов прогрессивной эволюции в мире животных.
Выяснение механизмов, препятствующих мутационному потоку в процессе индивидуального развития, и разработка вопроса о значении таких механизмов в прогрессивной эволюции -г это в действительности познание еще одного фактора стабильности в биологии. Жизнь на нашей планете представлена хорошо обособленными, дискретными формами, которые образуют собой видовую мозаику биосферы. Стабильность генома в ряду поколений обеспечивается механизмами наследственности. Имеющиеся мутационные нарушения генетического аппарата половых клеток незначительны, хотя и необходимы для эволюционного преобразования.
Стабильность индивидуального развития — залог осуществления наследственной преемственности. Иными словами, стабильность в процессе онтогенеза требуется для доставки наследственного материала от особей одного поколения особям другого. Од-
ним из механизмов генетической стабильности соматических клеток в процессе онтогенеза является специфический иммунитет. Это утверждение справедливо для большинства многоклеточных форм жизни. Без формирования в онтогенезе и эволюционного развития в филогенезе механизмов иммунологического контроля за соматическим мутагенезом жизнь достаточно организованных многоклеточных была бы невозможна.
Источник
Возникновение иммунитета
Эволюция формировала систему иммунитета около 500 млн. лет. Этот шедевр природы восхищает нас красотой гармонии и целесообразностью.
До середины 60-х годов 20 века считалось, что все беспозвоночные, включая хордовых, обладают неспецифической формой реагирования, а позвоночные, начиная с круглоротых, имеют также и специфический иммунитет.
Однако в опытах с трансплантацией аллогенной (чужеродной) ткани некоторым разновидностям червей и даже губок было показано, что пересаженная ткань может подвергаться специфическому отторжению с формированием кратковременной иммунологической памяти. Возможно, губки были первыми животными, у которых появился специфический трансплантационный иммунитет, так как у некоторых их разновидностей отмечается неспецифическое отторжение трансплантата без формирования иммунологической памяти.
Схема эволюции иммунитета
- 1. Первыми иммунными клетками были клетки, способные осуществлять фагоцитоз. Изначально их функцией было пищеварение, затем способность к фагоцитозу стала использоваться в борьбе с чужеродными агентами и собственными измененными клетками. Способность к фагоцитозу, появившись у одноклеточных, сохранилась и у млекопитающих.
- 2. Специфический клеточный иммунитет, вероятно, появился у некоторых разновидностей губок и кишечнополостных (например, у гидр, коралловых полип). Тогда же, возможно, появился предшественник лимфоцита, хотя иммунная клетка, имеющая маркер Т-лимфоцита, впервые обнаружена у кольчатых червей.
- 3. Система гистосовместимости, возможно, появилась тогда же, когда и трансплантационный иммунитет, то есть у губок. Подтверждением тому может служить то, что моноклональные антитела к одному из клонов в эксперименте подавляют реакцию отторжения трансплантата этого клона клеток. При изучении МНС классов I и II хрящевых рыб и позвоночных было обнаружено сходство в кодирующей их ДНК. Появление у рыб молекул двух классов гистосовместимости закрепилось в эволюции.
- 4. Синтез Ig — привилегия позвоночных, хотя иммуноглобулиноподобные молекулы были выявлены у мене развитых видов начиная с иглокожих (морских звезд, морских ежей и других).
- 5. Зачатки тимуса и костного мозга впервые появились у круглоротых, хотя прообразы иммунных органов появились уже у кольчатых.
Роль иммунитета в эволюции
Увеличение числа клеток в организме ведет за собой увеличение риска возникновения мутаций. Это является своеобразной платой за многоклеточность. Для контроля за мутационным потоком необходима развитая система иммунитета. Возможно, именно развитие многоклеточности явилось той посылкой, которая дала толчок к развитию иммунитета. По другой причине, мы получили такой сложный иммунитет в следствие сложного и долгого развития в течение всей жизни, а также по невозможности к большому числу репродукции. Поэтому наш организм защищает нас от всеразличных патогенов и антител для сохранения нашего существования.
Развитие иммунной системы шло параллельно с усложнением организации животного и освоением им новых мест обитания, где оно могло встретиться с новыми факторами.
Определенную роль играли увеличение продолжительности жизни и интенсификация обменных процессов. Например, маложивущим животным не нужна длительная иммунологическая память.[3]
Источник
Наука иммунология происходит от латинского слова immunitas (освобождение). Она изучает весь комплекс событий, которые приводят к уничтожению антигенно чужеродных веществ, с которыми приходит в контакт организм. Традиционное понимание иммунитета как способа защиты от инфекционных микроорганизмов (вирусов, бактерий, простейших) сегодня изменилось. Иммунные механизмы защиты проявляются всегда, когда конкретный организм сталкивается с теми или иными чужеродным в антигенном отношении материалом – будь то бактерии, вирусы, мутационно измененные собственные клетки тела, тканевые и органные трансплантаты или простые химические соединения, которым приданы иммунногенные свойства. В настоящее время иммунитет рассматривается как способ защиты организма от всех антигенно чужеродных веществ как экзогенной, так и эндогенной природы; биологический смысл подобной защиты – обеспечение генетической целостности особей вида в течение их индивидуальной жизни.
Таким образом, иммунитет выступает в качестве фактора стабильности онтогенеза (т.е. стабильности его гомеостаза). Необходимое условие передачи наследственного материала от поколения к поколению (осуществления генетического гомеостаза) – это сохранение уникального набора генов и их полноценной работы.
Два основных механизма иммунной защиты:
1. Неспецифический иммунитет – система защитных факторов организма, присущих данному виду как наследственно обусловленное свойство.
Включает: А) Барьерную функцию эпителия;
Б) Гуморальные факторы
ü лизоцим слюны и слизистых секретов, лейкоцитов – нейтрофилов, эозинофилов и макрофагов;
ü дефенсины фагоцитирующих лейкоцитов;
ü интерфероны вирус-инфицированных клеток;
ü система комплемента – сывороточные белки, неактивные «полуфабрикаты», конвертируемые в активную форму при контакте с антигеном (специфический путь), либо с другими патогенами (неспецифический путь);
В) Эндоцитоз (фаго — и пиноцитоз)
Г) Комплексная реакция воспаления (calor, dolor, rubor, tumor, functia lese) – повышение температуры, появление боли в очаге, покраснение, опухоль и нарушение функции. Способствует миграции фагоцитирующих лейкоцитов из сосудистого русла в очаг повреждения;
Д) Активность NK-клеток (натуральных «киллеров») – крупных гранулярных лимфоцитов без четкой Т- или В-дифференцировки, оказывающих прямое цитотоксическое на вирус-инфицированные, злокачественно трансформированные и патоген-содержащие (поглотившие патоген) клетки.
2. Специфический иммунитет – комплексная адаптационная (приобретенная) система реакций, характеризующихся высокой специфичностью в распознавании антигена, индуцибельностью и формированием иммунологической памяти.
Включает:
Центральные органы иммунной системы (ИС): тимус, костный мозг – места образования лимфоцитов – основных клеток, ответственных за специфический иммунитет и
Периферические органы ИС: селезенка, лимфатические узлы и сосуды, лимфоидная ткань ЖКТ (желудочно-кишечного тракта): пейеровы бляшки, аппендикс, миндалины; диффузно распределенные лимфоциты почек, печени и соединительной ткани – локусы миграции и локализации лимфоцитов из центральных органов.
Лимфоциты относятся к той категории клеток, которые широко распространены по организму. Различные типы организации лимфоцитов обеспечивают наиболее эффективную работу ИС при встрече с чужеродными антигенами. Клетки, принимающие участие в становлении и функционировании ИС можно разделить на две группы:
1) Т- и В-лимфоциты и их субпопуляции
2) Макрофаги.
Становление и роль иммунитета, рассмотренные в эволюционном аспекте, позволяют также понять, что прогресс в мире животных по линии увеличения абсолютного количества соматических клеток (от типа Губки и Типа Кишечнополостные к Типу Хордовые и его представителю – человеку) и обеспечивает эволюцию специфического иммунитета.
Неоценимый вклад в развитие эволюционной иммунологии вообще внесли исследование П. Медавара, продемонстрировавшие иммунологический конфликт как основную причину несовместимости трансплантата донора и организма реципиента и показавшие, что причина кроется в антигенных различиях между двумя организмами, заведомо обладающими генетической уникальностью и, следовательно, специфическими белковыми составами. Именно в недрах трансплантационной иммунологии выяснена роль тимуса в реакциях клеточного реагирования и окончательно сформировано представление о двух формах иммунного ответа – клеточного и гуморального.
М.Ф. Бернет (1964) первым определил иммунитет как реакцию организма, направленную на подержание генетической целостности организма (генетического гомеостаза). Действительно, рост интенсивности спонтанного мутагенеза в соматических клетках является неизбежным следствием возрастающего числа клеток у многоклеточных особей. Чем больше делящихся и дифференцирующихся соматических клеток в организме, тем выше вероятность генетических ошибок в их геноме и тем эффективнее должен быть контроль за мутационным процессом.
Цель: Изучить главные эволюционные пути становления отдельных форм иммунной системы от одноклеточных и первых многоклеточных до наиболее совершенных типов и классов животных.
Знать: Современное определение иммунитета, виды иммунитета, основные эволюционные этапы формирования иммунной защиты, основные звенья иммунной системы млекопитающих.
Уметь: Применить общий эволюционный принцип к пониманию становления эволюции иммунной защиты и многоклеточности.
Материалы и оборудование: рабочая тетрадь, ручка, простой карандаш, набор цветных карандашей, калькулятор.
Вопросы для самоподготовки:
1. Основные положения учения Ч. Дарвина.
2. Современное понятие «иммунитет».
3. Специфический и неспецифический (врожденный) иммунитет.
4. Клетки, такни и органы иммунной системы (костный мозг, тимус – центральные органы иммунитета; селезенка, лимф. узлы и др. периферические структуры иммунной системы
5. B- и T- системы иммунитета
6. Эволюция иммунной системы от т. Простейшие до т. Хордовые
7. Основные ароморфозы в эволюции иммунитета
8. Коэволюция специфического иммунитета и многоклеточности в филогенезе. Причина?
РАБОТА В АУДИТОРИИ
Работа № 1. Соотношение между различными формами иммунитета и уровнем организации в мире животных. (Рассмотреть и зарисовать схему-рисунок по В.Г. Галактионову, 2005 с некоторыми изменениями):
Работа № 2. Элементы неспецифического (врожденного) иммунитета:
А) Отметить барьерную функцию эпителия на конкретных примерах: Зарисовать таблицу и пояснения к ней.
Б) Перечислите гуморальные факторы неспецифического иммунитета
Таблица. Механизмы антибактериальной защиты, осуществляемые эпителием (по Галактионову В.Г., 2005).
| Фактор защиты | Эффекторы |
| Механический | Плотные межклеточные контакты, смыв микроорганизмов жидкостными секретами и пассажем воздуха вдоль эпителиальных покровов |
| Химический | Жирные кислоты кожи, лизоцим (слюна, слезы, пот), низкий pH (желудок); пепсин (кишечник), антибактериальные пептиды дефенсины (кишечник) |
| Микробиологический | Конкуренция с нормальной микрофлорой кишечника за источники питания и преимущественную колонизацию эпителия, продукция бактериями специфических химических соединений |
Работа № 3.Принципиальная схема развития воспалительного процесса:
Изучить и зарисовать таблицу и отметить, что характерные признаки комплексной воспалительной реакции: повышение температуры, появление боли в очаге, покраснение, опухоль и нарушение функции направлены на усиление кровотока, возрастание проницаемости и приток фагоцитарных клеток для осуществления фагоцитоза и активации цитокинов.
Работа № 4.Принципиальные механизмы поддержания генетического гомеостаза в животном мире.
Изучить с использованием материалов вышеуказанного методического пособия по теме занятия, существование трех звеньев защиты генетической целостности организма и описать их сущность: 1) внутриядерной системы репарации ДНК; 2) внутриклеточной системы самоуничтожения мутационно поврежденных клеток путем запуска механизма апоптоза; 3) иммунной системы защиты макроорганизма от потенциально опасных мутационно измененных клеток.
1) Два основных способа «починки» разрывов двойной спирали ДНК. Первый способ (негомологичное соединение концов) чреват неточностями — потерей или вставкой лишних нуклеотидов в районе разрыва. Второй более точен, но требует наличия «запасной копии» поврежденного фрагмента ДНК. В наших клетках в течение суток могут быть исправлены лишь несколько структурных изменений ДНК.
2) Апоптозом называют механизм программируемой и регулируемой гибели клеток. Механизм апоптоза включается, в частности, при повреждениях систем репарации ДНК и накоплении повреждений ДНК. Эти изменения активируют ряд специфических протеаз в клетке, которые, в свою очередь, активируют эндонуклеазы (разрывающие связи внутри НК).
3) Эффективность работы разнообразных звеньев Т — клеточного иммунитета – одно из важнейших условий защиты организма от злокачественных новообразований. На усиление этих звеньев направлена и специфическая терапия при онкологических заболеваниях. (источник рисунка: интернет-ресурс shvarz.livejournal.com/310996.html)
Работа № 5.Основы генетической уникальности индивидуума (иммуногенетики) и проблемы трансплантационного иммунитета.
На примере главного генетического комплекса гистосовестимости HLA (human leukocyte antigen) показать высокую степень генетической и белковой уникальности индивидуумов в человеческой популяции.
Антигены генетического комплекса HLA 6-й хромосомы кариотипа человека играют главную роль в тканевой совместимости донора и реципиента. Локус ABC комплекса HLA имеет протяженность 2 морганиды и содержит 15 аллельных вариантов гена А, 20 аллелей гена B и 5 аллелей гена С. Учитывая затруднительность расчета возможных форм взаимодействия этих генов и, следовательно, фенотипов, возможно, тем не менее, установить число уникальных вариантов генотипов. Рассчитайте приблизительное число лиц с одинаковыми генотипами по локусу АВС в г. Тюмени (население около 600 000 чел.), в России (около 140 млн. человек) и на планете Земля (около 6 млрд. человек), если всего существует 120 комбинаций генотипов по аллелю А, 210 – по аллелю В и 15 – по аллелю С.
Алгоритм действий:
1) Рассчитать вероятность полного совпадения генетического полиморфизма у двух человек по локусам А,B и C, используя закон умножения вероятностей:
1/120 x1/210 x 1/15 = 1/378000.
2) Рассчитать приблизительное число лиц с одинаковыми генотипами по г. Тюмени, России и Земному шару.
ЗАНЯТИЕ 2.6.3.
Источник