Индуцированный иммунитет у растений
ОСНОВЫ ИММУНИТЕТА РАСТЕНИЙ К БОЛЕЗНИ
При самой суровой эпифитотии растения поражаются болезнью неодинаково, что связано с устойчивостью и иммунитетом растений. Под иммунитетом понимается абсолютная непоражаемость при наличии инфекции в условиях, благоприятных для заражения растений и развития болезней. Устойчивость — это свойство организма противостоять сильному поражению болезнями. Эти два свойства зачастую отождествляют, имея в виду слабое поражение растений болезнями.
Устойчивость и иммунитет — сложные динамичные состояния, которые зависят от особенностей растения, возбудителя болезни и условий внешней среды. Изучение причин и закономерностей устойчивости очень актуально, так как только в этом случае возможна успешная работа по выведению устойчивых сортов.
Категории иммунитета
Иммунитет бывает врожденным (наследственным) и приобретенным. Врожденный иммунитет передается от родителей потомству. Он изменяется только с изменением генотипа растения.
Приобретенный иммунитет формируется в процессе онтогенеза, что достаточно часто встречается в медицинской практике. У растений такого четко выраженного приобретенного свойства нет, но существуют приемы, позволяющие повысить устойчивость растений к болезням. Они активно изучаются.
Пассивная устойчивость определяется конституциональными особенностями растения, независимо от действия патогена. Нанример, толщина кутикулы некоторых органов растений является фактором пассивного иммунитета. Факторы активного иммунитета действуют только при контакте растения и возбудителя, т.е. возникают (индуцируются) в период патологического процесса.
Выделяют понятие специфического и неспецифического иммунитета. Неспецифический — это неспособность некоторых возбудителей вызвать заражение определенного вида растений. Например, свекла не поражается возбудителями головневых заболеваний зерновых культур, фитофторозом картофеля, картофель не поражается церкоспоро-зом свеклы, зерновые — макроспориозом картофеля и т. д. Иммунитет, проявляющийся на уровне сорта по отношению к специализированным возбудителям, называется специфическим.
Установлено, что устойчивость определяется суммарным действием защитных факторов на всех этапах патологического процесса. Все многообразие защитных факторов подразделяется на 2 группы: препятствующие внедрению патогена в растение (аксения); препятствующие распространению патогена в тканях растений (истинная устойчивость).
В первую группу входят факторы или механизмы морфологического, анатомического и физиологического характера.
Анатомо-морфологические факторы. Преградой для внедрения возбудителей может служить толщина покровных тканей, строение устьиц, опушенность листьев, восковой налет, особенности строения органов растений. Толщина покровных тканей является защитным фактором в отношении тех возбудителей, которые проникают в растения непосредственно через эти ткани. Это в первую очередь мучнисто-росяные грибы и некоторые представители класса Оомицеты. Строение устьиц имеет значение для внедрения в ткань бактерий, возбудителей ложных мучнистых рос, ржавчин и др. Обычно через плотно прикрывающиеся устьица возбудителю внедриться труднее. Опушенность листьев защищает растения от вирусных болезней, насекомых, передающих вирусную инфекцию. Благодаря восковому налету на листьях, плодах и стеблях капли на них не задерживаются, что препятствует прорастанию грибных патогенов.
Габитус растений и форма листьев также являются факторами, препятствующими начальным стадиям заражения. Так, сорта картофеля с рыхлым строением куста меньше поражаются фитофторозом, так как лучше проветриваются и инфекционные капли на листьях высыхают быстрее. На узкие листовые пластинки оседает меньше спор.
Роль строения органов растений можно проиллюстрировать на примере цветков ржи и пшеницы. Рожь очень сильно поражается спорыньей, в то время как пшеница — очень редко. Это объясняется тем, что у цветков пшеницы цветковые чешуи не раскрываются и споры возбудителя практически не проникают в них. Открытый тип цветения у ржи не препятствует попаданию спор.
Физиологические факторы. Быстрому внедрению возбудителей может препятствовать высокое осмотическое давление в клетках растений, скорость физиологических процессов, приводящих к затягиванию ран (образование раневой перидермы), через которые проникают многие патогены. Важна также скорость прохождения отдельных фаз онтогенеза. Так, возбудитель твердой головни пшеницы внедряется только в молодые проростки, поэтому сорта, дружно и быстро прорастающие, поражаются меньше.
К механическим преградам относятся клетки с толстыми оболочками или клетки, у которых в стенках откладывается лигнин — вещество, не поддающееся разрушению ферментами паразитов.
Отсутствие (или недостаток) в растительных тканях веществ, необходимых для развития патогена. Любая растительная ткань представляет собой питательный субстрат, на котором патоген способен паразитировать. Обычно наиболее сильно поражаются хорошо обводненные ткани, богатые растворимыми углеводами и аминокислотами. На определенных этапах онтогенеза, когда какое-либо вещество еще не синтезировано растением или оно уже претерпело изменения в процессе метаболизма, устойчивость к заболеваниям выше. Так, гриб Fu-sarium graminearum Schw. паразитирует на зерновых только при наличии в тканях таких сложных органических соединений, как холин и бетаин. Их больше всего содержится в пыльниках, поэтому колос поражается фузариозом после фазы цветения.
Ингибиторы. Это соединения, содержащиеся в растительных тканях или синтезированные в ответ на заражение, которые подавляют развитие патогенов. К ним относятся фитонциды — вещества различной химической природы, являющиеся факторами врожденного пассивного иммунитета. В большом количестве фитонциды вырабатываются тканями лука, чеснока, черемухи, эвкалипта, лимона и др.
Алкалоиды — азотсодержащие органические основания, образующиеся в растениях. Особенно богаты ими растения семейства бобовых, маковых, пасленовых, астровых и др. Например, соланин картофеля и томатин помидоров токсичны для многих возбудителей. Так, развитие грибов рода Fusarium тормозится соланином в разведении 1:105. Подавлять развитие возбудителей могут фенолы, эфирные масла и ряд других соединений. Все перечисленные группы ингибиторов всегда присутствуют в интактных (неповрежденных тканях).
Индуцированные вещества, которые синтезируются растением в процессе развития патогена, называют фитоалексинами. По химическому составу все они — низкомолекулярные вещества, многие из них
имеют фенольную природу. Установлено, что сверхчувствительная реакция растения на заражение зависит от скорости индукции фитоалексинов. Известны и идентифицированы многие фитоалексины. Так, из растений картофеля, зараженных возбудителем фитофтороза, выделены ришитин, любимин, фитуберин, из гороха — пизатин, из моркови — изокумарин. Образование фитоалексинов представляет типичный пример активного иммунитета.
К активному иммунитету относится также активизация ферментных систем растения, в частности окислительных (пероксидаза, поли-фенолоксидаза). Это свойство позволяет инактивировать гидролитические ферменты возбудителя болезни и обезвреживать им токсины.
Приобретенный, или индуцированный, иммунитет. Для повышения устойчивости растений к инфекционным болезням применяется биологическая и химическая иммунизация растений.
Биологическая иммунизация достигается обработкой растений ослабленными культурами патогенов или продуктами их жизнедеятельности (вакцинация). Ее применяют при защите растений от некоторых вирусных болезней, а также бактериальных и грибных патогенов.
Химическая иммунизация основана на действии некоторых химических веществ, в том числе и пестицидов. Ассимилируясь в растениях, они изменяют обмен веществ в направлении, неблагоприятном для возбудителей болезней. Примером таких химических иммунизаторов служат фенольные соединения: гидрохинон, пирогаллол, ортонитрофенол, паранитрофенол, которыми обрабатывают семена или молодые растения. Иммунизирующим свойством обладает ряд фунгицидов системного действия. Так, дихлорциклопропан защищает рис от пирикуляриоза благодаря усилению синтеза фенолов и образованию лигнина.
Известна иммунизирующая роль и некоторых микроэлементов, входящих в состав ферментов растений. Кроме того, микроэлементы улучшают поступление основных элементов питания, что благоприятно сказывается на устойчивости растений к болезням.
Генетика устойчивости и патогенности. Типы устойчивости
Устойчивость растений и патогенность микроорганизмов, как и все другие свойства живых организмов, контролируются генами, одним или несколькими, качественно отличающимися друг от друга. Наличие таких генов обусловливает абсолютный иммунитет к определенным расам патогена. Возбудители болезни, в свою очередь, имеют ген (или гены) вирулентности, позволяющий ему преодолевать защитное действие генов устойчивости. По теории X. Флора, на каждый ген устойчивости растения может выработаться соответствующий ген вирулентности. Это явление называют комплементарностью. При воздействии патогена, обладающего комплементарным геном вирулентности, растение становится восприимчивым. Если гены устойчивости и вирулентности некомплементарны, клетки растения локализуют возбудитель в результате сверхчувствительной реакции на него.
Например (табл. 4), согласно этой теории, сорта картофеля, имеющие ген устойчивости R,, поражаются только расой 1 возбудителя P. infestans или более сложной, но обладающей обязательно геном вирулентности 1 (1,2; 1,3; 1,4; 1,2,3) и т. д. Сорта, не имеющие генов устойчивости (г), поражаются всеми без исключения расами, в том числе и расой без генов вирулентности (0).
Гены устойчивости чаще всего доминантны, поэтому их сравнительно легко передать потомству при селекции. Гены сверхчувствительности, или R-гены, определяют сверхчувствительный тип устойчивости, которую называют также олигогенной, моногенной, истинной, вертикальной. Она обеспечивает растению абсолютную непоражаемость при воздействии на него рас без комплементарных генов вирулентности. Однако с появлением в популяции более вирулентных рас патогена устойчивость теряется.
Другой тип устойчивости — полигенная, полевая, относительная, горизонтальная, которая зависит от совокупного действия множества генов. Полигенная устойчивость в различной степени присуща каждому растению. При высоком ее уровне патологический процесс замедляется, что дает возможность растению расти и развиваться, несмотря на пораженность болезнью. Как любой полигенный признак, подобная устойчивость может колебаться под воздействием условий выращивания (уровень и качество минерального питания, влагообеспеченность, длина дня и ряд других факторов).
Полигенный тип устойчивости наследуется трансгрессивно, поэтому закрепить его путем селекции сортов проблематично.
Распространенным является сочетание сверхчувствительной и по-лигенной устойчивости в одном сорте. В этом случае сорт будет иммунным до появления рас, способных преодолевать моногенную устойчивость, после чего защитные функции определяет полигенная устойчивость.
Методы создания устойчивых сортов
В практике наиболее широко используются направленная гибридизация и отбор.
Гибридизация. Передача генов устойчивости от род ительских растений потомству происходит при межсортовой, межвидовой и межродовой гибридизации. Для этого в качестве родительских форм подбирают растения с желаемыми хозяйственно-биологическими характеристиками и растения, обладающие устойчивостью. Донорами устойчивости чаще бывают дикие виды, поэтому в потомстве могут появиться нежелательные свойства, которые устраняются при возвратных скрещиваниях, или беккроссах. Бейер ос сы повторяют до тех пор, пока все признаки <<дикаря», кроме устойчивости, не поглотятся сортом.
С помощью межсортовой и межвидовой гибридизации создано много сортов зерновых, зернобобовых культур, картофеля, подсолнечника, льна и других культур, устойчивых к наиболее вредоносным и опасным болезням.
При нескрещиваемости некоторых видов друг с другом прибегают к методу «посредника», при котором каждый вид родительских форм или один из них скрещивают сначала с третьим видом, а затем полученные гибриды скрещивают между собой или с одним из первоначально планируемых видов.
В любом случае устойчивость гибридов проверяют на жестком инфекционном фоне (естественном или искусственном), т. е. при большом количестве инфекции возбудителя, в условиях, благоприятных для развития болезни. Для дальнейшего размножения отбирают растения, сочетающие высокую устойчивость и хозяйственно ценные признаки.
Отбор. Этот прием — обязательный этап при любой гибридизации, но он может быть и самостоятельным методом получения устойчивых сортов. Методом постепенного отбора в каждом поколении растений с нужными признаками (в том числе и с устойчивостью) получено много сортов сельскохозяйственных растений. Он особенно эффективен для перекрестноопыляющихся растений, так как потомство их представлено гетерозиготной популяцией.
С целью создания устойчивых к болезням сортов все более широко применяются искусственный мутагенез, генная инженерия и др.
Причины потери устойчивости
Со временем сорта, как правило, утрачивают устойчивость либо в результате изменения патогенных свойств возбудителей инфекционных болезней, либо нарушения иммунологических свойств растений в процессе их воспроизводства. У сортов со сверхчувствительным типом устойчивости она теряется с появлением более вирулентных рас или комплементарных генов. Сорта с моногенной устойчивостью поражаются из-за постепенного накопления новых рас патогена. Вот почему селекция сортов только со сверхчувствительным типом устойчивости является бесперспективной.
Причин, способствующих образованию новых рас, несколько. Первая и наиболее частая — мутации. Они обычно проходят спонтанно под действием различных мутагенных факторов и присущи фитопатогенным грибам, бактериям и вирусам, причем для последних мутации — единственный способ изменчивости. Вторая причина — гибридизация генетически разных особей микроорганизмов при половом процессе. Этот путь характерен главным образом для грибов. Третий путь — гетерокариоз, или разноядерность, гаплоидных клеток. У грибов разноядерность может возникать из-за мутаций отдельных ядер, перехода ядер из разнокачественных гиф по анастомозам (сросшимся участкам гиф) и перекомбинации генов при слиянии ядер и последующем их делении (парасексуальный процесс). Разноядерность и пар асексуальный процесс имеют особенное значение для представителей класса несовершенных грибов, у которых отсутствует половой процесс.
У бактерий, помимо мутаций, существует трансформация, при которой ДНК, выделенная одним штаммом бактерий, поглощается клетками другого штамма и включается в их геном. При трансдукции отдельные сегменты хромосомы из одной бактерии переносятся в другую с помощью бактериофага (вируса бактерии).
У микроорганизмов образование рас идет постоянно. Многие из них сразу же погибают, будучи неконкурентоспособными из-за более низкого уровня агрессивности или отсутствия других важных признаков. Закрепляются в популяции, как правило, более вирулентные расы при наличии сортов и видов растений с генами устойчивости к существующим расам. В таких случаях новая раса даже при слабой агрессивности, не встречая конкуренции, постепенно накапливается и распространяется.
Например, при возделывании картофеля с генотипами устойчивости R,, R4 и R1R4 в популяции возбудителя фитофтороза будут преобладать расы 1; 4 и 1,4. При введении в производство сортов с генотипом R2 вместо R4 из популяции патогена постепенно исчезнет раса 4, а распространятся расы 2; 1,2; 1,2,4.
Иммунологические изменения сортов могут происходить и в связи с изменением условий их произрастания. Поэтому перед районированием сортов с полигенной устойчивостью в других эколого-географи-ческих зонах обязательно проводят их иммунологическое испытание в зоне будущего районирования.
5.3. Приобретенный (индуцированный) иммунитет
Успехи, достигнутые в области медицинской иммунологии, послужили стимулом к поиску аналогичных явлений в фитоиммунологии. Исследовали природу индуцированной устойчивости у растений и возможности ее практического применения. Но когда в них не обнаружили антител и стало ясно глубокое отличие в природе иммунитета животных и растений, изучение этого вопроса затормозилось. Однако в последние 20 лет интерес к проблеме приобретенного (индуцированного) иммунитета вновь повысился. Это было связано с несколькими причинами. 1) Устойчивость выведенных селекционных сортов растений недолговечна и рано или поздно они начинают поражаться вновь появившимися расами паразитов. 2) Химическая защита растений довольно дорога, а грибы приспосабливаются к фунгицидам, которые становятся неэффективными. Кроме того, часто неконтролируемое и безграмотное использование фунгицидов в больших количествах загрязняет окружающую среду и вредно отзывается на здоровье животных и человека. 3) Есть возможность регулировать синтез защитных веществ у восприимчивых растений внешними воздействиями.
Устойчивость растений к заражению может повышаться как в результате перенесенного заболевания, так и в результате вакцинации.
О механизме повышения устойчивости растений после перенесенного вирусного заболевания мы уже говорили. Разберем теперь приемы вакцинации.
В настоящее время широко используется вакцинация ослабленными штаммами вирусов. Наибольшие успехи достигнуты при вакцинации томатов вирусом табачной мозаики для борьбы с мозаичным заболеванием томатов в условиях закрытого грунта. Опрыскивание растений ослабленной культурой ВТМ повышает урожай и не сказывается на качестве плодов. Такого рода вакцинация успешно осуществляется в Голландии, Швеции, Дании, Англии, Канаде, США, Японии, а также в нашей стране.
Имеются работы, указывающие на возможность вакцинации и других культур. Так, слабопатогенный штамм ВТМ S7 использовался для защиты томатов от столбура (микоплазмы), огурца — от вируса зеленой крапчатой мозаики. Показано, что картофель, вакцинированный вирусом X, не заражался фитофторозом, а также другими вирусами. Защитные свойства вакцинации сохранялись при этом в течение ряда лет.
Этот метод, однако, должен использоваться с осторожностью, поскольку никогда нельзя исключить появления в популяции вакцинного штамма агрессивных форм, которые поразят устойчивые сорта. Поэтому он используется пока лишь в условиях теплиц.
Вакцинацию растений можно осуществлять также патогенными грибами. Так, для иммунизации огурцов использовали ослабленные культуры (сохранявшиеся в чистой культуре 14 лет) листового и стеблевого штаммов возбудителя аскохитоза Ascochyta cucumeris. Эти штаммы задерживали проявление симптомов и развитие заболевания, в то время как непатогенные штаммы гриба не обладали вакцинирующим действием.
В других опытах первый настоящий лист растений огурца заражали патогеном Colletotrichnm lagenarium, вызывающим антракноз. Через 4 дня у организма развивалась системная устойчивость, т. е. все выше расположенные, а также не раскрывшиеся листья становились устойчивыми к заражению этим и другими паразитами. При этом обнаружена лигнификация тканей прямо под аппрессорием патогена. В другом случае использовали для иммунизации растений горшечного табака конидии Peronospora tabacina, которые помещались в почву вокруг стебля. В листьях развивалась системная устойчивость. Эти приемы вакцинации, как и в случае с вирусами, также должны использоваться с осторожностью, поскольку не исключена возможность заражения растений.
В качестве вакцины в настоящее время используются также индукторы паразита. Например, из мицелия грибов Phytophthora infestans и Fusarium culmorum выделен липогликопротеидный комплекс (ЛГП) — биогенный индуктор, который в концентрации 0,0005% вызывает повышение устойчивости картофеля, томатов и сахарной свеклы к различным болезням. Он обладает системным действием: при обработке клубней картофеля и семян томатов и сахарной свеклы повышенную устойчивость к болезням приобретали растения, выросшие в полевых условиях. Их урожайность повышалась на 10 — 30%. Действие этого индуктора основано не на фунгитоксичности, а на повышении естественного свойства устойчивости. Предполагается, что одним из механизмов действия индуктора является вызываемая им системная сенсибилизация растений. Под влиянием сенсибилизирующих доз ЛГП клетки клубня картофеля и других растений приобретают отдельные черты ультраструктуры, характерные для специализированных терпеноидогенных клеток, с которыми связан биосинтез терпеноидов и фенолов. А фитоалексины картофеля и томатов, как мы уже отмечали выше, принадлежат к числу сесквитерненоидов. Сенсибилизированные клетки, таким образом, уже подготовлены для массового синтеза фитоалексинов, которые при встрече с патогеном быстро образуются и накапливаются.
Таким образом, сенсибилизация связана с функциональной перестройкой клеток клубня в сторону повышения их биосинтетических возможностей, и в первую очередь, с образованием специализированных защитных веществ: терненоидных фитоалексинов, суберина. Возрастают при этом и энергетические возможности сенсибилизированных клеток (увеличивается число митохондрий, исчезает крахмал из амилопластов, используемый в качестве энергетического субстрата).
Основным недостатком этого приема иммунизации растений является дороговизна получения индуктора. Поэтому перед исследователями стоит задача совершенствования технологии их производства.
Химическая иммунизация. Этот прием вакцинации подразумевает обработку растений химическими веществами, которые регулируют протекание защитных реакций. Технологически этот метод прост (растения опрыскивают растворами веществ или замачивают в них семена), перспективен, поскольку можно получить много соединений, влияющих на метаболизм и не обладающих токсическим действием на человека и окружающую среду.
В качестве препаратов-иммунизаторов используют стимуляторы роста, фенолы, макро- и микроэлементы и пр. Они обладают системным действием, и, не оказывая влияния на патоген, изменяют метаболизм растения-хозяина, повышая его устойчивость. Их называют иногда веществами внутри растительного действия.
Каковы механизмы повышения устойчивости растений под влиянием химических иммунизаторов? Например, при опрыскивании водной эмульсией полиакриловой кислоты на поверхности растения образуется пленка, которая препятствует проникновению ростков ржавчины; ауксины повышают устойчивость клеточной стенки к пектолитическим ферментам патогена; гербициды диаллат на пшенице и дихлорциклопропан на рисе способствуют активизации фенольного метаболизма и окислительных процессов, усиливают синтез лигнина и способствуют увеличению прочности клеточных стенок; такие соединения, как антибиотик хлорамфеникол, гербицид симазин, ретарданты индуцируют синтез фитоалексинов; гербициды, ретарданты обладают сенсибилизирующим действием, т. е. подготавливают растение к заражению.
В последние годы в связи с достижениями в области биотехнологии для повышения устойчивости растений к заболеваниям начали использовать методы клеточной инженерии. В частности, для получения растений, устойчивых к грибам или фитонатогенным бактериям, используется следующая последовательность приемов, представленная на схеме (рис. 20). Получают растения-регенеранты, проверяют их на устойчивость и выделяют линии, которые значительно превосходят исходный сорт по устойчивости к бактерии или грибу.
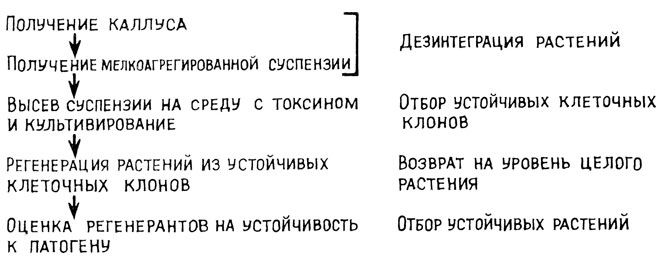
Рис. 20. Схема получения устойчивых к патогенам растений методом клеточной инженерии
Широко используется также метод верхушечной меристемы для оздоровления растений от вирусной инфекции. Для этого выделяют меристему, свободную от вируса, получают из нее каллусную ткань, а затем растение-регенерант, свободное от вируса, которое можно высаживать в почву.
* * *
Таким образом, в основе фитоиммунитета лежит множество защитных реакций. Но все они отражают уже завершающую фазу взаимоотношений двух партнеров, конечный неспецифический ответ клетки на контакт с несовместимым патогеном. Между тем, будет ли растение устойчиво или восприимчиво, т. е. индуцирование защитных реакций должно осуществляться на начальных этапах взаимодействия с патогенами, когда происходит распознавание партнеров. Именно распознавание «своего» от «чужого» является первичным процессом, тогда как ответные защитные реакции развиваются спустя какое-то время, необходимое для реализации процесса узнавания.
Различают 2 фазы в процессах, развивающихся после встречи растения и патогена: детерминантная — партнеры распознают друг друга, в результате чего определяется, будут ли они совместимыми или нет; экспрессивная — выражает события, продиктованные детерминантной фазой, — быстрое или медленное включение защитных реакций.
Какие продукты распознает растение? Это метаболиты паразита, называемые индукторами или элициторами. Они находятся на поверхности патогена или в составе его выделений. Очевидно, растение в процессе эволюции «научилось» распознавать такие метаболиты паразита и использовать их как триггер для ответного включения всего комплекса защитных реакций. Большинство индукторов представляет собой высокомолекулярные соединения, в составе которых имеются углеводные группы. В очень низких концентрациях (10-5 M) они вызывают некротическую реакцию (реакцию сверхчувствительности) и накопление фитоалексинов.
Хорошо изученным индуктором является липогликонротеидный комплекс, выделенный из клеточного содержимого гриба возбудителя фитофтороза картофеля Phytophthora infestans. О химической природе других индукторов мы уже говорили в разделе о фитоалексинах.
Механизм распознавания до настоящего времени экспериментально еще не изучен. Согласно гипотезе Альберсхейма и Андерсон-Проути [11], индукторы патогена представляют собой как бы антигенную детерминанту для их распознавания рецепторными белками (лектинами) в составе мембран растения-хозяина. Продуктом гена авирулентности патогена должен быть фермент, ответственный за синтез молекулы индуктора, тогда как продуктом гена устойчивости является система, участвующая в образовании рецепторного белка па мембране растения-хозяина. Поскольку ген устойчивости растения и ген авирулентности паразита комплементарны, то происходит взаимодействие их продуктов, что и приводит к включению реакции сверхчувствительности и отторжению паразита.
Считается, что в процессе распознавания вполне вероятен межклеточный контакт партнеров посредством лектин-углеводного взаимодействия. Имеются наблюдения, подтверждающие возможность такого механизма взаимодействия растения-хозяина и патогена.
Так, обнаружено, что гликопептиды клеточной поверхности гриба Phytophthora infestans, содержащие глюкозамин — специфический лиганд для лектина картофеля, — связываются с плазмалеммой, выделенной из паренхимных клеток клубней картофеля. При этом в устойчивой комбинации это связывание сильнее, чем в неустойчивой. Факт взаимодействия плазмалеммы с гликоконъюгатами гриба подтверждается также подавлением гемагглютинирующей активности плазмалеммы гликопептидами патогена.
В плазменной мембране сои обнаружены сайты со способностью активно связывать глюканы клеточной стенки патогенного гриба Phytophthora megasperma. Эти сайты могут играть роль в элицитор-обусловленном защитном ответе. Белок мембран, связывающий глюкан, сохраняет свою связывающую активность после солюбилизации. Гликопротеины из клеточной стенки уредоспор Puccinia graminis tritici селективно связывались с лектино-подобными белками, выделенными из листьев пшеницы.
О действии индукторов (элициторов) на уровне поверхности клетки свидетельствуют также наблюдения, в которых показано, что элициторы ряда патогенов обладают способностью деполяризовать плазмалемму клеток растений. Этот эффект был быстрым, стабильным и обратимым. Обратимость действия элицитора свидетельствует о том, что он не повреждает растительную мембрану.
Большинство известных в настоящее время индукторов являются неспецифическими, т. е. они вызывают защитные реакции у всех сортов растений (устойчивых и неустойчивых к патогену, из которого они выделены) и присутствуют у всех рас гриба. Именно неспецифические индукторы распознаются растением и, очевидно, такой механизм лежит в основе видового иммунитета.
А какова же природа специфической, сортовой устойчивости? Каким образом специфические паразиты препятствуют включению защитных реакций?
По мнению большинства исследователей, специфические патогены научились подавлять неспецифический защитный ответ растений. Так, из мицелия гриба Phytophthora infestans были выделены вещества, которые подавляли реакцию сверхчувствительности у картофеля, вызываемую инфицированием несовместимой расой. Эти вещества, получившие название супрессоров (фактор совместимости), представляют собой низкомолекулярные глюканы со связями β-1,3- и β-1,6- и содержатся лишь в совместимых расах. Эти соединения облегчают распространение несовместимого гриба но тканям растений, индуцируя в них восприимчивость. К числу супрессоров, кроме токсических глюканов, относятся также токсины паразитов и импедины (нетоксичный бактериальный фактор подавляющий защитный механизм растения-хозяина).
Как же действуют супрессоры? Предполагается, что в совместимой комбинации супрессоры, выделяемые патогеном, конкурируют с индуктором за рецепторный участок на плазмалемме растения-хозяина, имея к нему большее сродство, и препятствуют таким путем включению защитных реакций.
Считают, что супрессор имеет 2 активных центра: неспецифическую группировку, которая конкурирует с индуктором за рецепторный участок у хозяина, и специфическую группу, которая изменчива. Поскольку растение в процессе эволюции приобретает новые гены сортовой устойчивости (R), предполагается, что продукты этих новых генов устойчивости связывают супрессор паразита по его вариабельной группировке и как бы «уводят» его с поверхности взаимодействия паразита и хозяина. Индуктор остается вне конкуренции и вновь взаимодействует с рецепторами, индуцируя защитные реакции. Такое растение приобретает сортовую устойчивость к патогену.
В результате мутаций гриба вариабельная часть супрессора (которая связывалась продуктом R-гена) изменяется так, что теряет комплементарность к этому продукту и перестает им связываться. Супрессор вновь оказывается на свободе и начинает вытеснять индуктор в борьбе за рецептор. Так возникает ген вирулентности паразита, способный преодолевать ген устойчивости. Далее процесс повторяется.
Встает вопрос: как же реализуется (или передается) сигнал, ведущий от распознавания к включению защитных реакций, т. е. от детерминантной к экспрессивной фазе? Ответ на этот вопрос в настоящее время отсутствует. В соответствии с гипотезой индуктор — рецептор вслед за связыванием индукторов несовместимой расы патогена рецептором растения через плазмодесмы между плазматической мембраной и ядрами контактирующих клеток и соседних клеток хозяина проходят вторичные мессенджеры, аналогичные циклическим нуклеотидам других организмов. Такие мессенджеры запускают процессы обмена и, в конечном счете, приводят к синтезу и накоплению фитоалексинов и запуску других защитных реакций.
В качестве вторичных мессенджеров могут функционировать эндогенные индукторы. Образование этих индукторов происходит в результате деятельности синтезируемой фитопатогенными грибами эндонолигалактуроназы, которая вызывает распад клеточных стенок растения-хозяина с образованием пектиновых фрагментов (олигосахаридов), действующих как эндогенные индукторы фитоалексинов. В настоящее время эти соединения привлекают внимание исследователей как возможные посредники ответа растения на стресс. Они индуцируют такие естественные защитные механизмы растений, как синтез фитоалексинов, лигнина, ингибиторов протеиназ, богатых гидроксипролином гликопротеинов. Способность олигосахаридов индуцировать защитные реакции растений является одним из аспектов общего свойства этих молекул регулировать генную экспрессию.
Роль возможного вторичного мессенджера могут выполнять также ионы Ca2+. В связи с ролью Ca2+ в преобразовании внешних воздействий в клеточный ответ с помощью кальмодулина — рецептора, образующего интегральную часть Ca2+-мессенджер системы, изучали роль Ca2+ в ускорении синтеза фитоалексинов (ришитина и любимина). Обнаружено, что при обработке тканей клубня картофеля арахидоновой кислотой — индуктором фитоалексинов — ионы Ca2+ стимулировали накопление ришитина и любимина. Добавление ЭГТА, которая связывает Ca2+ клубней, подавляло накопление фитоалексинов. Эти наблюдения позволяют предположить, что ионы Ca2+ каким-то путем могут регулировать первичные процессы взаимодействия паразита с растением-хозяином — процессы распознавания, включающие затем защитную реакцию, т. е. Ca2+ выступает как вторичные мессенджеры.