У кого иммунитет к малярии

Восприимчивость к малярии практически всеобщая.
В эндемичных районах новорожденные, родившиеся от иммунных матерей, обладают пассивным иммунитетом до 2-3 месяцев. К шестимесячному возрасту защитные антитела в крови уже не обнаруживаются.
Имеются популяции населения, которые обладают полной или частичной врожденной невосприимчивостью к определенным видам возбудителей.
Африканские негры обладают врожденной, генетически обусловленной невосприимчивостью к P.vivax, которая до настоящего времени сохраняется у их потомков — американских негров. Причина невосприимчивости заключается в недостаточности у негроидов антигенов крови группы Даффи (Fy или Fy), которые являются паразитарными рецепторами, факторами прикрепления паразита к эритроцитам.
Выраженной устойчивостью к P.falciparum обладают носители гемоглобина S (HbS) и других патологических глобинов — С и В. По-видимому, малярийный паразит не способен усваивать аномальные гемоглобины. Установлено, что у неиммунных лиц с HbS при заражении P.falciparum тропическая малярия протекает с невысокой паразитемией и значительно легче, чем у больных с нормальным гемоглобином, а при наличии приобретенного иммунитета наблюдается полная невосприимчивость.
Имеется корреляция в распространении талассемии и малярии. Относительная устойчивость к малярии гетерозиготных носителей талассемии объясняется недостатком в эритроцитах необходимого паразиту железа.
У лиц с дефицитом глюкозо-6-фосфат-дегидрогеназы также имеется невосприимчивость к Р falciparum.
Приобретенный иммунитет при малярии включает клеточный и гуморальный ответы. Началом иммунных процессов, которые стимулируют выработку антител, является фагоцитирование малярийных паразитов макрофагами, в цитоплазме которого происходит процессинг поглощенного белка с выделением иммуногенных детерминант или э пито по в. Макрофаг через интерлейкин-1 (ИЛ-1) стимулирует Т-лимфоцит хелперного ряда, который помогает «загустить» иммунный ответ. Антигены предъявляются лимфоцитам Т-хелперам вместе с комплексом гистосовместимости (HLA), презентация способствует выроботке интерлейкина-2 (ИЛ-2), который и активизирует размножение и дифференцировку различных клонов В-клеток, синтезирующих специфические антитела. Антитела атакуют эпитоны антигенов, нарушая их функцию. Кроме того, ИЛ-2 стимулирует размножение и дифференцировку Т-цитотоксических лимфоцитов, которые атакуют инфицированные эритроциты в непосредственном клеточном контакте. Антигенными раздражителями в естественных условиях являются только эритроцитарные стадии плазмодиев. Антигенными детерминантами, которыми обладают не только поверхностные белки различных стадий развития паразита, но и его функциональные белки, в частности протеин-р41, обладающий в плазмодии альдолазной активностью. Он способствует выработке энергии, которая используется на репликацию и созревание плазмодия в эритроците. Спорозоиты, гаметоциты и тканевые стадии паразитов не вызывают активности иммунного ответа и, следовательно, образования к ним специфических антител.
Развивающийся иммунитет обуславливает синхронизацию размножения паразитов в эритроцитах с упорядочением лихорадочных пароксизмов, постепенное снижение паразитемии и в дальнейшем спонтанное прекращение приступов.
Образование специфических антител начинается с первых дней заболевания. Серологические реакции становятся положительными через 5-12 дней от начала заболевания. Наиболее высокие титры отмечаются через 4-6 недель. Затем наблюдается снижение титров антител, но серологические реакции остаются положительными в низких титрах еще в течение двух лет при тропической малярии, 6 лет — при трехдневной и 15 лет — при четырехдневной малярии.
У лиц, живущих в эндемичных районах, в результате постоянного ре инфицирования развивается высокий иммунитет, поэтому клинические проявления малярии выражены слабо, либо отсутствуют совсем. Но прекращение повторных заражений, в случае выезда из эндемичной зоны, приводит к постепенному угасанию иммунитета и при заражении малярией спустя несколько лет, повлечет за собой тяжелое течение заболевания.
Как и при других инфекциях, иммунитет при малярии строго в ид о- и штаммоспецифичен. Установлено, что быстрее вырабатывается иммунитет к P.vivax и P.malariae и медленнее — к P.falciparum. При обследовании населения деревень Центральной Африки получены следующие данные: в первые годы жизни дети заражаются всеми видами возбудителей малярии и тяжело болеют. К 9 годам у них из крови исчезает P.vivax, к 14 годам — P.malariae, и только после 16 лет вырабатывается некоторый иммунитет к тропической малярии. Иммунитет при овале-малярии развивается быстро, о чем свидетельствует быстрое спонтанное прекращение приступов при этой инфекции.
Известно, что размножение малярийных плазмодиев приводит к истощению иммунной системы инфицированного человека, что может частично объяснить развитие на фоне тропической малярии лимфомы Беркита, Показано, что инфицирование малярией приводит к иммунодефициту со снижением количества Т-хелперов, запускающих иммунный ответ, так как именно Т-хелперы элиминируются при малярийных приступах (Лалаянц И.Э., Милованова Л.С. 1988).
С 70-х годов проводятся исследования по созданию эффективных вакцин, однако к настоящему времени вакцин, пригодных для человека, нет. В то же время новейшие молекулярно-биологические исследования позволяют глубже проникнуть в биологию малярийного плазмодия и генез иммунного ответа, проследить динамику смены антигенов различных стадий развития плазмодия, их функциональную структуру. Можно надеяться, что в скором будущем будут созданы долгожданные действенные вакцины против малярии.
Читайте также:
Малярия | Симптомы малярии | Лечение малярии | География распространения малярии | Возбудитель малярии | Малярия в истории | Патогенез при малярии | Эпидемиология малярии
Иммунитет при малярии. Признаки и клиника малярииУ больных малярией иммунитет возникает под влиянием антигенного воздействия эритроцитарных стадий развития паразита и паразитемии, при которых в самом начале болезни наблюдается повышение активности клеточных субпопуляций Т-лимфоцитов, Т-хелперов и Т-супрессоров. Благодаря им после приступов малярии постепенно снижается интенсивность паразитемии. Иммунитет при малярии (как и при туберкулезе) нестерильный, напряженность его со временем снижается, что является одной из причин наступления ранних рецидивов болезни, которые наблюдаются при всех формах малярии, при трехдневной и овале-малярии возможны и отдаленные рецидивы вследствие активизации гипнозоитов, находящихся в клетках печени. Иммунитет после перенесенной малярии видовоспецифический, кратковременный и недостаточно стойкий. У коренных жителей эндемичных по малярии местностей она протекает чаще всего в виде легких форм или в форме паразитоносительства. При выезде из этих местностей иммунитет утрачивается и они становятся, как и все другие люди, поголовно восприимчивыми к малярии. Некоторые группы людей — коренные жители Западной Африки и негритянское население Северной Америки — имеют генетически обусловленный врожденный естественный иммунитет к малярии, вызываемой PI. vivax. К другим видам малярии, они так же как и все люди, восприимчивы. Такая индивидуальная устойчивость к малярии vivax обусловлена недостаточностью у них изоантигенов в эритроцитах, поверхность которых не содержит рецепторов для Pi. vivax. Относительная резистентность к тропической малярии у некоторых коренных жителей обусловлена наличием аномального гемоглобина S(HbS). У лиц с дефицитом Г-6-ФДГ малярия протекает легче и с невысокой паразитемией. В эндемичных местностях новорожденные дети, как правило, не болеют малярией благодаря пассивному иммунитету, полученному от матери (A.M. Бронштейн, А.К. Токмалаев, 2002 г.)
Яркая клиническая картина малярии обусловлена триадой симптомов: озноб, жар, потоотделение, последовательная смена которых характеризует малярийный приступ (пароксизм). Приступы малярии наблюдаются при всех ее видах с некоторыми клиническими нюансами. Различают четыре клинические формы малярии: трехдневную, тропическую, четырехдневную и овале-малярию. Минимальный инкубационный период при тропической малярии длится 8-14 дней, при четырехдневной — 3-6 недель. При трехдневной малярии, вызываемой PI. vivax (южный штамм), он составляет 7-21 день, при северном штамме (PI. vivax hibemans)-6-14 мес(иногда до одного года и более). При трехдневной овале-малярии его продолжительность — 11-16 дней. У лиц, систематически принимавших с целью профилактики противомалярийные препараты, инкубационный период может удлиняться (B.C. Матковский, А.Я. Лысенко, К.М. Лобан и др.). Клинические проявления малярии зависят от вида возбудителя, тяжести течения и формы болезни. Наиболее характерным признаком малярии является смена периодов лихорадочных приступов (пароксизмов) и нормальной температуры (апирексии), что характерно для интермиттирующего типа лихорадки. Первое повышение температуры происходит обычно остро среди полного здоровья. Острому началу болезни может предшествовать непродолжительный (1-2 дня, реже 3-5 дней) продромальный период, который чаще отмечается при первичной инфекции и проявляется общим недомоганием, мышечной слабостью, разбитостью, познабливанием, ломотой в мышцах и суставах, снижением аппетита, тошнотой, сонливостью, головной болью, субфебрильной температурой. Затем наступает типичный пароксизм, при котором озноб, как правило, носит потрясающий характер; больной вынужден лечь в постель, старается тепло укрыться, чтобы согреться. У него стучат зубы, лицо и конечности становятся бледно-синюшными, холодными, хотя температура тела быстро повышается и достигает высоких цифр (39-40°С и выше). Фаза озноба, длящегося от 30-60 мин до 2-3 ч, сопровождается болью в пояснице, мышечной дрожью, иногда рвотой; пульс учащен, дыхание поверхностное. К концу этого периода больной согревается, лицо краснеет, кожа становится горячей, сухой, усиливается головная боль. Наступает вторая фаза малярийного приступа — период жара, длительность которого при трехдневной малярии составляет 6-12 ч, при четырехдневной — 4-5 ч, а при тропической растягивается на 30-40 ч. В этом периоде головная боль усиливается, отмечаются спутанность сознания, бред, возбуждение, одышка, иногда судороги, снижается АД. Через несколько часов жар сменяется фазой обильного потоотделения, температура быстро снижается или резко падает до нормы. Обессиленный после приступа больной засыпает. Общая продолжительность малярийного приступа 6-12 ч, при тропической малярии -до суток и более. Далее наступает период апирексии, в течение 24 или 48 ч в зависимости от продолжительности эритроцитарной шизогонии у разных видов возбудителей. — Также рекомендуем «Клиника малярийного приступа. Анемия при малярии» Оглавление темы «Тропические инфекционные болезни»: |
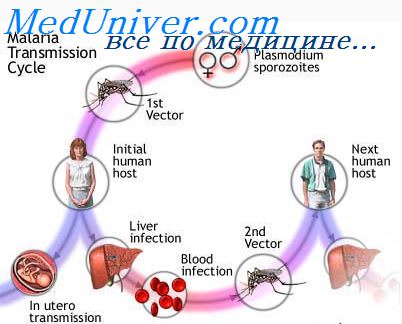
О различиях в восприимчивости к заражению возудителем указано выше, здесь же следует отметить, что механизмы восприимчивости определяются многочисленными факторами, в том числе морфологическими особенностями передней кишки комара, скоростью и объемом поглощаемой инфицированной крови, набором аминокислот в гемоцеле и др. Установлено, что заражение плазмодием влияет на продолжительность жизни комара, способность его к полету и, возможно, другие физиологические качества имаго.
Численность популяции переносчика определяется природными факторами: площадью мест выплода, продолжительностью развития личиночной стадии, продолжительностью гонотрофического цикла и жизни имаго. Масштабы выплода зависят от количества и ритма выпадения осадков, структуры почвы, температуры и относительной влажности воздуха и типа водоемов — мест выплода, которые предпочитает конкретный вид переносчика. Например, большое количество осадков приводит к увеличению площади мест выплода An. culicifacies и к уменьшению — An. fluviatilis.
Значительное снижение или повышение температуры воздуха приводит к резкому снижению численности популяции и сезонному перерыву передачи. На территориях с умеренным климатом зимовка переносчика может происходить на стадии имаго в состоянии диапаузы или на стадии личинки. Па участках ареала с субтропическим, тропическим и экваториальным климатом во время сухого и жаркого сезона выплод переносчиков продолжается в незначительных масштабах, а у самок .наблюдается феномен эстивации — медленное переваривание крови во время длительного нахождения на дневках с подходящим микроклиматом.
Редким, но тем не менее значимым является другой механизм передачи при гемотрансфузии от донора-паразитоносителя реципиенту или при групповом использовании непромытых шприцев, что часто наблюдается у наркоманов. Гемотрансфузионным путем наиболее часто передается P. malariae, бессимптомное носительство которого может продолжаться десятки лет. В гиперэндемичных очагах у рожденных от иммунных матерей вследствие разрыва плаценты и попадания паразитов в организм новорожденного наблюдаются случаи врожденной малярии.
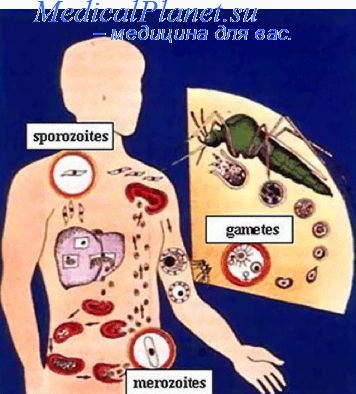
Восприимчивость населения к малярии не является всеобщей. Что касается врожденной невосприимчивости, то в настоящее время накоплены значительные данные относительно ее механизмов на эритроцитарных стадиях развития возбудителя. Указанные механизмы связаны с особенностями структуры мембраны, а также внутренней среды эритроцита и имеют генетическую основу. Одним из примеров врожденной невосприимчивости к малярии является невосприимчивость представителей негроидной расы Западной Африки к заражению P. vivax.
Было установлено, что в мембране эритроцитов у представителей этой расы отсутствуют изоантигены FyFy, что препятствует проникновению паразита в эритроцит и тем самым — дальнейшему развитию инвазии. Гетерогенность популяций эритроцитов человека по таким признакам, как вариант гемоглобина и изоантигены, представленные в гомозиготной или гетерозиготной формах, влияют в различной степени на восприимчивость к малярии. Наиболее изученным является вопрос о влиянии HbS на течение малярии.
У гомозигот HbS наблюдается серповидно-клеточная анемия, что приводит к смерти в детском или подростковом возрасте. У гетерозигот HbS отмечаются относительно доброкачественное течение тропической малярии, умеренная паразитемия, низкая смертность, что, по-видимому, объясняется удалением из кровотока пораженных эритроцитов до завершения в них шизогонии. Выявлено совпадение ареалов тропической малярии и группы населения с высокой частотой HbS. Аналогичные данные получены в отношении носительства HbC, HbE, a также HbF — фетального гемоглобина, который обеспечивает определенную степень защиты новорожденных от тропической малярии, однако механизмы невосприимчивости остаются невыясненными.
Дефицит глюкозо-6-фосфат-дегидрогеназы (Г-6-Ф-Д) — одного из ферментов эритроцита, распространенный в странах Средиземноморья, а также в Азербайджане, способствует смягчению клинических проявлений болезни, в частности, вызванных P.vivax, и более длительному паразитоносительству, что имеет существенное эпидемиологическое значение.
Приобретенный иммунитет к малярии формируется в результате предшествующего заражения, ограничивает или предотвращает развитие клинических форм и патологических процессов после реинвазии и является строго видо- и штаммоспецифичным. Пассивный иммунитет имеют новорожденные от гипериммунных матерей, получившие антитела класса IgG внутриутробно и после рождения — с молоком матери. Длительность постинвазионного иммунитета различная, чаще до 1 года, однако при новом заражении протективный иммунитет формируется быстрее, чем при первой встрече с возбудителем. Продолжительность циркуляции сывороточных противомалярийных антител классов IgG и IgM более длительная, однако их титры резко падают в течение 6-9 мес после лечения инвазированных.
Механизмы иммунного ответа сложны и включают как клеточный, так и гуморальный компоненты, причем клеточный компонент имеет более выраженную протективную значимость. Мощность иммунной прослойки в населении прямо пропорциональна интенсивности передачи. В очагах с очень интенсивной передачей протективный иммунитет формируется к 6-10 годам и поддерживается в более старших возрастных группах за счет суперинвазирования. Однако, несмотря на высокую толерантность к клиническим проявлениям инфекции, развитие возбудителя в организме продолжается, и более старшие возрастные группы являются, как и дети, эффективными источниками возбудителя инфекции.
В очагах с менее интенсивной передачей формирования выраженного иммунного ответа наблюдается в более старших возрастных группах (20-30 лет и старше), острые клинические проявления наблюдаются не только у детей, но и у подростков и взрослых. Па участках ареала с низкой интенсивностью передачи коллективный иммунитет населения выражен слабо, тем не менее за счет иммунной памяти длительность периода клинических проявлений у взрослых короче, чем у детского населения.
До настоящего времени средства специфической профилактики малярии не разработаны. Интенсивные исследования по созданию противомалярийной вакцины под эгидой ВОЗ проводятся в трех основных направлениях: разработка антиспорозоитной, антимерозоитной и антигаметной вакцины с использованием при этом методов генной инженерии. Прототипы антиспорозоитной и антигаметной вакцины, созданные в конце 80-х годов, оказались недостаточно иммуногенными, поэтому перспективы получения и использования в практике борьбы с малярией эффективной вакцины в течение ближайших 5-10 лет остаются проблематичными.
— Читать далее «Восприимчивость к малярии. Очаги малярии»
Оглавление темы «Гепатиты С и Д. Эпидемиология малярии»:
1. Вирусный гепатит С. Распространенность вирусного гепатита С
2. Механизм передачи гепатита С. Эпидемиологический процесс гепатита С
3. Гепатит Д. Возбудитель гепатита Д
4. Эпидемиологический процесс при гепатите Д. Профилактика гепатита Д
5. Малярия. Распространенность малярии
6. Возбудители малярии. Гаметоцитогония при малярии
7. Возбудитель тропической малярии. Особенности возбудителей малярии
8. Источник малярии. Механизм передачи малярии
9. Восприимчивость к малярии. Иммунитет при малярии
10. Восприимчивость к малярии. Очаги малярии