Взаимодействие клеток при гуморальном иммунитете

Глава 11. Взаимодействие клеток при гуморальном иммунном ответе
ВЗАИМОДЕЙСТВИЕ В- и Т-КЛЕТОК
Т- и В-лимфоциты распознают разные участки молекулы антигена
Большой вклад в изучение различных функций Т- и В-клеток внесли исследования, проведенные в конце 1960 — начале 1970 гг. Митчисоном и другими исследователями с использованием химически модифицированных белков. Ими было установлено, что для индукции оптимального вторичного ответа с образованием антител против небольшом молекулы, или гаптена (он обладает иммуногенностью только в том случае, когда связан с белком-носителем), животное необходимо иммунизировать не гаптеном в свободной форме, а конъюгатом гаптен-носитель, и затем повторно ввести тот же конъюгат. Этот феномен был назван эффектом носителя. Исследуя различные клеточные популяции, авторы этих работ обнаружили, что за распознавание носителя ответственны Т-клетки, тогда как гаптен распознают В-лимфоциты (рис. 11.6). Дальнейшие эксперименты были направлены на подробное изучение того, каким образом В-клеточные антитела-рецепторы распознают эпитопы, а Т-клетки — фрагменты процессированного антигена.

Рис. 11.6. Облученным рентгеновскими лучами мышам трех групп были введены примированные антигеном клетки селезенки и после этого различные антигены. 1. В группе 1 (спленоциты, примированные ДНФ-БСА, + конъюгат ДНФ-БСА) мыши продуцировали большое количество антител к ДНФ. 2. В группе 2 (спленоциты, примированные ДНФ-БСА, + ДНФ-ОА) образование антител к ДНФ было слабым (эффект носителя отсутствовал). 3. В группе 3 (спленоциты, примированные ДНФ-БСА, + спленоциты, примированные ОА, + ДНФ-ОА) наблюдалась сильная реакция образования антител к ДНФ. Этот опыт показывает, что для продукции антител к гаптену нет необходимости в примировании носителем: достаточно использовать примированные носителем клетки селезенки. (ДНФ — динитрофенол; БСА — бычий сывороточный альбумин; ОА — овальбумин.)
Одним из принципиально важных выводов в этих исследованиях был следующий: индивидуальная В-клетка может получать «помошь» от Т-клеток, специфичных к различным антигенным пептидам, при условии что она способна презентировать их детерминанты каждой Т-клетке. В ходе иммунною ответа in vivo взаимодействие с В-лимфоцитами (вызывающее их пролиферацию и дифференцировку) осуществляют, как считается, Т-клетки, уже стимулированные в результате предшествующего контакта с антигеном, презентированным другими АПК (например, дендритными клетками). На этих данных основаны представления о клеточных взаимодействиях при анти гелообразовании (рис. 11.7). Согласно им, антиген, попавший в организм, процессируется клетками, которые презентируют его в высокоиммуногенной форме Тх-клеткам и В-клеткам. Детерминанты антигена, распознаваемые Т-лимфоцитами, отличаются от детерминант, распознаваемых В-клетками, которые делятся и дифференцируются в антителообразующие. Таким образом, для активации В-клетки необходимы два следующих процесса:
• взаимодействие «нативного» антигена с Ig-peцепторами В-клетки;
• стимуляция сигналом (или сигналами) от Тх-лимфоцитов, отвечающих на связанный с молекулами МНС процессированный антиген.

Рис. 11.7. АПК, например дендритные клетки, презентируют антиген непримированным Т-клеткам. В-лимфоциты также связывают антиген и презентируют его Т-клеткам, получая от них сигнал к делению и дифференцировке в антителообразующие клетки (АОК) и В-клетки иммунологической памяти (Вп).
Взаимодействие В- и Т-клеток происходит с участием костимулирующих молекул
Путем выращивания и клонирования Т-клеток в присутствии антигена, АПК и ИЛ-2 можно получить популяцию антигенспецифичных Т-лимфоцитов. Это позволяет выявить in vitroнепосредственно взаимодействующие молекулярные кластеры на поверхности Т и В-клеток. В такой экспериментальной системе было установлено, что при взаимодействии происходит поляризация Т-клеток с концентрированием ТкР на стороне, обращенной к В-клеткам. В свою очередь В-клетки также поляризуются и экспрессируют большую часть своих антигенов МНС класса II и ICAM-I в непосредственной близости к Т-клеткам. Взаимодействие между этими кластерами, очевидно, предполагает интенсивный обмен информацией, следствием которого становятся два важных события в жизненном цикле В-клеток: индукция пролиферации и дифференцировка в антителообразующие клетки.
Взаимодействие между Т- и В-клетками представляет собой двунаправленный процесс: В-клетки презентируют антиген Т-клеткам и в свою очередь получают от них сигналы к делению и дифференцировке. Центральное, специфическое взаимодействие происходит при этом между комплексом молекула МНС класса II—антиген и Т-клеточным рецептором; оно усиливается за счет связывания LFA-3 с CD2, а также ICAM-1 или IСАМ-3 с LFA-1 (рис. 11.8). В процессе участвуют и другие молекулы клеточной поверхности: В7-1 и В7-2 на В-клетках взаимодействуют с CD28, что ведет к стабилизации мРНК для ИЛ-2 и других цитокинов в Т клетках и в результате к продлению периода, в течение которого Т-клетка генерирует сигналы активации.

Рис. 11.8. Мембраносвязанный иммуноглобулин (mlg) взаимодействует с антигеном (Аг) и переводит его во внутриклеточный компартмент, где антиген подвергается деградации и образующиеся пептиды могут образовать комплекс с молекулами МНС класса II. Другие стрелки показывают установленные отдельные пути передачи сигнала. А и Б — передача сигналов, возникающих при взаимодействии антигена с рецепторами; она включает фосфорилирование тирозина и расщепление фосфоинозитида. Антигенные рецепторы регулируют также аффинность LFA-1 к ICAM-1, возможно посредством передачи сигнала. Т-клеточная молекула CD28 передает Т-клетке особый сигнал (В). Наиболее сильный активирующий сигнал для В-клеток возникает при стимуляции CD40 (Г). Кроме того, молекулы МНС класса II, по-видимому, индуцируют отдельные сигнальные процессы (Д). На рисунке не показан межклеточный обмен растворимыми цитокинами и их связывание соответствующими рецепторами клеток. (С разрешения по A. DeFranco, 199! Nature 351: 603-5.)
К настоящему времени установлено, что наиболее мощный сигнал для активации В-клеток обеспечивает молекула CD40; этот сигнал даже более эффективен, чем опосредуемые поверхностными иммуноглобулинами. В процессе этой активации Т-клетки временно экспрессируют взаимодействующий с CD40 лиганд — CD40L. Белок CD40 принадлежит к семейству рецепторов ФНО, a CD40L является отдаленно родственным представителем этого семейства. Их взаимодействие способствует вступлению В-лимфоцитов в клеточный цикл. Данный сигнал (связывание CD40 с CD40L) имеет важное значение для формирования центров размножения и развития гуморального иммунного ответа на Т-зависимые антигены. Подтверждением этого служит развитие иммунодефицитного заболевания — синдрома гиперпродукции IgM — в результате мутации гена CD40L. Данное расстройство характеризуется тем, что Т-клетки не оказывают «помощи» В-лимфоцитам и в результате образуются только низкоаффинные антитела IgM и отсутствует переключение синтеза иммуноглобулинов на продукцию высокоаффинных IgG или IgA.
При взаимодействии Т- и В-клеток высвобождение Т-клеточных цитокинов (например, ИЛ-4) поляризовано в направлении В-клеток. Синтезируемые некоторыми В-лимфоцитами ИЛ-1 и ИЛ-6 повышают экспрессию ИЛ-2 Т-клетками. Поскольку, однако, лишь некоторые В-клетки секретируют эти цитокины, большинство В-клеток способно, по-видимому, активировать только ранее примированные Т-лимфоциты или Т-клетки памяти.
Не так давно было установлено, что Т-клетки CD4+у мыши и человека можно разделить на две субпопуляции в зависимости от профиля продуцируемых ими цитокинов (см. гл. 10).
• Т-лимфоциты CD4+, образующие ИЛ-2 и ИФγ, но не ИЛ-4 (они обозначены Tx1), ответственны главным образом за реакции гиперчувствительности замедленного типа, однако у мыши они могут также «помогать» В-клеткам продуцировать IgG2α и в значительно меньшей степени IgGI или IgE.
• Т-клетки CD4+, образующие ИЛ-4 и ИЛ-5, но не ИЛ-2 или ИФγ, отнесены к другой субпопуляции, Тх2. У мыши эти клетки служат весьма эффективными хелперами в продукции антител, в основном IgGI и IgE.
• Многие Т-клетки, главным образом у человека, занимают промежуточное положение по типу образуемых ими цитокинов и названы «Тх0»-клетками. Однако доказано и присутствие у человека классических Tx1- и Тх2-клеток, наиболее характерное для патологически измененных тканей.
Секреция и действие Т-клеточных цитокинов важны для активации В-клеток
При взаимодействии Т- и В-клеток Т-лимфоциты могут секретировать ряд цитокинов, оказывающих сильное действие на В-клетки. Такими цитокинами служат ИЛ-2 (индуктор пролиферации Т- и В-клеток), ИЛ-4 (действует на ранней стадии активации и пролиферации В-клеток), ИЛ-5 (мощный активатор В-клеток у мыши, но не у человека) и ИЛ-6 (эффективный стимулятор В-клеточной дифференцировки) (рис. 11.9). Кроме того, Т-клетки синтезируют ФНОα и β. Имеются сообщения о том, что эти молекулы важны для роста В клеток. Активацию В-лимфоцитов вызывают и другие цитокины, в частности ИЛ-11 и ИЛ-13, по характеру действия сходный с ИЛ-4. Более подробно роль цитокинов в активации Т- и В-клеток рассмотрена ниже.

Рис. 11.9. В-клетки активируются антигеном (Аг), представленным на антигенпрезентирующих клетках (АПК), например макрофагах, в присутствии ИЛ-4 и ИЛ-1. Это индуцирует экспрессию рецепторов для ИЛ-2 и других цитокинов. ИЛ-2, ИЛ-4 и ИЛ-5 (у мыши) вызывают деление клеток. На рисунке показан только один цикл клеточного деления, хотя обычно клетки делятся многократно. На дифференцировку лимфоцитов в антителообразующие клетки (АОК) влияют ИЛ-4, ИЛ-5 (у мыши), ИЛ-6, ИЛ-10 и ИФγ. (Ат – антитело.)
Результатом взаимодействия Т- и В-лимфоцитов может быть либо активация, либо инактивация (анергия)
Приведенные выше данные о взаимодействии Т- и В клеток указывают на то, что единственно возможный его результат — активация В-лимфоцитов. Однако в действительности это не так. Как было отмечено выше, кооперация АПК с Т-клетками способна давать диаметрально противоположные результаты, а именно вызывать либо активацию, либо инактивацию (клональную анергию) клеток. То же самое имеет место и в случае В лимфоцитов, которые часто переходят в состояние анергии. Этот процесс очень важен в связи с тем, что созревание аффинности антител в холе иммунного ответа — результат гипермутирования генов, кодирующих вариабельные области антител, — может легко приводить к образованию высокоаффинных аутоантител. Клональная анергия и другие формы периферической толерантности необходимы для того, чтобы такие потенциально опасные клоны «молчали». Однако на молекулярном уровне этот процесс еще мало изучен. Кроме того, остается неизвестным значение IgM и IgD, двух поверхностных антигенспецифичных рецепторов В-клеток, в процессах активации и инактивации. По-видимому, в обоих случаях IgM- и IgD-рецепторы способны передавать соответствующие сигналы.
Для стимуляции В-клеток Т-независимыми антигенами не требуется помощи Т-клеток
В иммунном ответе на большинство антигенов участвуют и Т-, и В-клетки, распознающие антиген сопряженно. Такие антигены называют Т-зависимыми. Однако некоторые антигены способны активировать В-клетки без помощи Т-клеток — это Т-независимые антигены. Для этих антигенов характерен ряд общих свойств (рис. 11.10). Так, все они представляют собой крупные полимерные молекулы с повторяющимися антигенными детерминантами. Многие из Т-независимых антигенов в высоких концентрациях обладают способностью активировать клоны В-клеток, специфичных к другим антигенам (феномен поликлональной В-клеточной активации). Однако в низких дозах они активируют только В-клетки соответствующей специфичности. Нередко Т-независимые антигены обнаруживают повышенную устойчивость к деградации. Это относится в основном к микробным антигенам, например бактериальным углеводам, таким как декстран и леван, а также к белкам бактерий, например флагеллину и эндотоксину.
Рис. 11.10. Основные свойства некоторых из наиболее распространенных Т-независимых антигенов. Эти антигены индуцируют образование цитокинов ИЛ-1, ФНО и ИЛ-6 макрофагами. (Следует указать, что поли-L-аминокислоты и мономерный бактериальный флагеллин являются Т-зависимыми антигенами, что свидетельствует о значении структуры антигена для его свойств (Т-зависимость или Т-независимость).
Первичный гуморальный отвез на Т-независимые антигены обычно немного слабее, чем на Т-зависимые, и достигает пика несколько раньше. В обоих случаях продуцируются главным образом антитела IgM. Однако вторичные иммунные ответы на антигены этих двух типов резко различаются. При повторном введении Т-независимого антигена ответ напоминает первичный, тогда как вторичная реакция на Т-зависимые антигены гораздо сильнее первичной и большую часть образующихся антител составляет IgG (рис. 11.11). Таким образом, Т-независимые антигены, по-видимому, не индуцируют, как правило, изменений, которые могли бы привести к созреванию ответа, характерному для случая Т-зависимых антигенов, которые вызывают переключение изотипа на продукцию IgG и повышение аффинности антител. Причиной этих различии между типами антигенов может быть отсутствие активации с участием CD40 в случае Т-независимых антигенов. Формирование иммунологической памяти при их воздействии также относительно слабое.
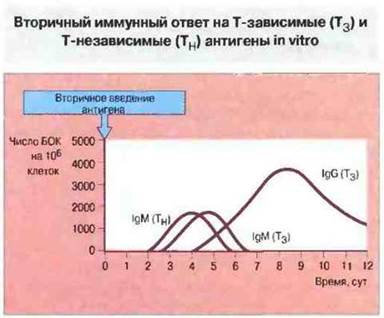
Рис. 11.11. Вторичный иммунный ответ на Т-зависимые антигены сильнее, чем первичный, что можно определить по накоплению большего числа продуцирующих IgG-антитела бляшкообразующих клеток (БОК) (см. гл. 29).
Механизм активации В-клеток Т-независимыми антигенами без участия Тх-клеток еще не раскрыт полностью. Возможно, полимерная структура этих антигенов позволяет им перекрестно связывать В-клеточные рецепторы, чему способствует устойчивость молекул антигена к деградации. Такой иммунный ответ, не требующий сложных клеточных взаимодействий, обеспечивает преимущество для выживания, поскольку позволяет организму быстрее реагировать на микробные антигены. Многие бактериальные антигены действуют независимо от Т-клеточной помощи, поскольку являются чрезвычайно сильными индукторами синтеза цитокинов ИЛ-1, ИЛ-6 и ФНОα макрофагами. Непродолжительность иммунной реакции на Т-независимые антигены и то, что она не включает образование IgG, может быть результатом отсутствия костимуляции, опосредованной CD40L, и отсутствия продукции ИЛ-2, ИЛ-4 и ИЛ-5, которые Т-клетки секретируют в реакции на Т-зависимые антигены. Часто пол влиянием Т-независимых антигенов активируются главным образом В-клетки субпопуляции В1, экспрессирующие маркер CD5. Эти клетки обнаруживаются преимущественно в брюшной полости.
Макрофаги участвуют в иммунном ответе на всех его этапах (рис. 4.9). Во-первых, они осуществляют немедленную защитную реакцию до тех пор, пока не произойдет усиление иммунного ответа, регулируемое антигенспецифичными Т-клетками. Во-вторых, они вызывают активацию Т-клеток, осуществляя процессинг и презентацию им антигена.

Рис. 4.9. Роль макрофага в иммунном ответе
Макрофаги осуществляют первоначальную защитную реакцию на ранней стадии ответа на инфекцию, до вступления в действие специфических механизмов иммунитета, зависимых от Т- и В-клеток. Позже функция макрофагов сводится к переработке {процессингу) и представлению {презентации) антигена. Наконец, распознавшие антиген Т-клетки выделяют цитокины, активирующие эффекторные фукции макрофагов.
Активированные, в свою очередь, Т-клетками, они выполняют важные функции в эффекторных механизмах клеточного иммунитета, вызывая воспаление и уничтожая микроорганизмы, а также опухолевые клетки.
Гуморальный иммунный ответ (образование антител) представляет собой ряд клеточных и молекулярных взаимодействий, происходящих в определенной последовательности:
- • АПК представляют Т-клеткам антиген, который последние распознают, переходя в активированное состояние;
- • Th-клетки и В-клетки взаимодействуют в результате презентации им антигенных фрагментов;
- • активированные В-лимфоциты пролиферируют и дифференцируются в антителообразующие клетки;
- • начинается синтез антител.
Презентация антигена Т-клетками. Процессинг антигена. Лишь очень небольшая доля молекул (
Проникшие в организм антигены подвергаются внутриклеточному процессингу—расщеплению на пептидные фрагменты, которые затем связываются с молекулами МНС. Эти фрагменты определяют антиген — специфичную активацию Т-клеток: рецепторы Т-клеток распознают аминокислотные последовательности этих фрагментов, связываемых в полости молекул МНС (рис. 4.10).
Взаимодействие между Т-лимфоцитами и клетками, составляющими гетерогенную группу так называемых «антигенпрезентиру- ющих клеток», — это наиболее детально изученный пример клеточной кооперации в иммунной системе. Взаимодействие Т-клеток и АПК после введения антигена открывает всю последовательность дальнейших событий и в основном определяет их конечный результат: если активируется достаточное количество хелперных Т-клеток (Th) CD4+, то почти всегда происходит активация В-клеток или развитие реакций клеточного иммунитета; если же стимуляция Th-клеток от- 70
сутстует, то может возникнуть та или иная форма иммунологической толерантности.

Рис. 4.10. Образование пептидных фрагментов и формирование комплекса антигена с МНС
Существуют различные типы АПК. Презентировать антигены могут самые разнообразные клетки — все зависит от того, как и где происходит первичное взаимодействие антигена с иммунной системой. Наиболее эффективно начальную активацию покоящихся Т-клеток CD4+ обеспечивают интердигитатные дендритные клетки (ИДК), присутствующие в изобилии в Т-клеточных зонах лимфатических узлов и селезенки. Для ИД К характерен высокий уровень экспрессии МНС-антигенов, которые взаимодействуют с Т-клеточным рецептором (ТкР) и молекулой CD4 на поверхности Th CD4+.
Интердигитатные клетки — это главный тип антигенпрезентирую- щих клеток, действующих при первичном иммунном ответе, поскольку они индуцируют пролиферацию Т-клеток эффективнее АПК всех других типов.
Именно клеточная пролиферация служит ключевым этапом развития иммунного ответа, обеспечивая увеличение количества ан- тигенспецифичных Т-клеток. Однако это лишь одна сторона эффективной активации Т-лимфоцитов. Способностью индуцировать и пролиферацию и хелперную функцию Т-клеток обладают также моноциты крови.
В качестве АПК могут действовать и В-клетки — они способны связывать, интернализовать и расщеплять специфический антиген на пептиды, которые образуют комплекс с молекулами МНС.
Презентацию антигена Т-клеткам обеспечивает взаимодействие множества молекул клеточной поверхности. Т-клеточный рецептор (ТкР) — димер, состоящий из a-цепи и p-цепи. Он распознает специфический пептид, находящийся в пептидсвязывающей полости молекулы МНС. Это связывание является определяющим для иммунологической специфичности, так как пептид, ассоциированный с МНС-молекулой, образует уникальную структуру, распознаваемую ТкР.